Úvod
Zmysly nám umožňujú prijímať podnety. Rozdeľujú sa na (obr. 1):
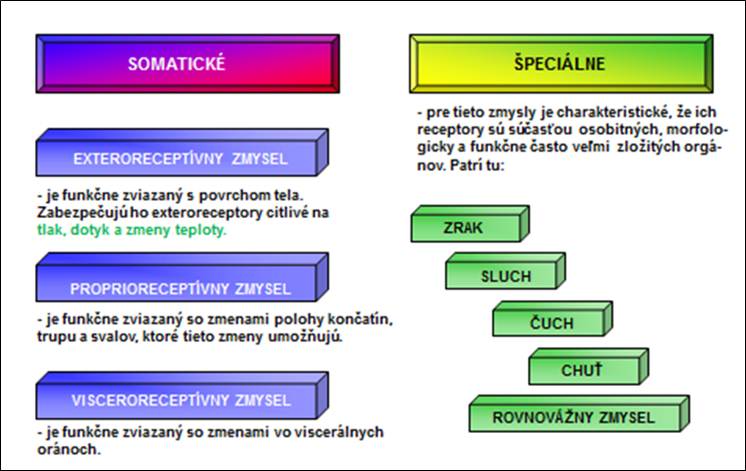
Obr. 1 . Rozdelenie zmyslov.
Zmyslové receptory
Sú voľné nervové zakončenia aferentných neurónov alebo tvarovo a funkčne špecializované bunky, ktoré zachytávajú podnety a zároveň kódujú ich intenzitu. Delia sa na
- Exoreceptory – prijímajú podnety z vonkajšieho prostredia;
- Endoreceptory – prijímajů podnety z vnútorneho prostredia organizmu.
Typy receptorov:
- Jednoduché receptory sú neuróny s nervovými zakončeniami
- Komplexné nervové receptory majú nervové zakončenia uzatvorené v dotykových tkanivových kapsuliach
- Väčšina špeciálnych senzorických receptorov sú bunky, ktoré uvoľňujú neurotransmiter do senzorických neurónov, vyvolávajúc tak akčný potenciál
Receptory poznáme:
- Tónické receptory – sú to pomaly adaptujúce sa receptory, u ktorých je po prvej aktivácii výboj rýchly, potom sa spomaľuje a nakoniec ich výbojová aktivita pretrváva tak dlho, ako pôsobí podnet.
- Fázické receptory – sú to rýchlo adaptujúce sa receptory čo znamená, že vykazujú výbojovú aktivitu po obdržaní podnetu, ale ak podnet pretrváva s konštantným pôsobením naďalej, ich výbojová aktivita sa zastavuje.
Klasifikácia receptorov:
- Mechanoreceptory reagujú na podnety mechanického charakteru. Poznáme:
- Baroreceptory – registrujú zmeny krvného tlaku;
- Rozťahové receptory pľúc – zachytávajú úroveň plnenia pľúc vzduchom pri vdychu;
- Proprioreceptory – reagujú na predĺženie svaloch a šliach;
- Taktilné receptory – reagujú na dotyk, tlak a vibrácie.
- Termoreceptory sú citlivé na zmenu teploty kože a delia sa na:
- Chladové receptory;
- Tepelné receptory.
- Chemoreceptory špecializujú sa na detekciu koncentrácií chemických látok v tekutinách alebo vo vzduchu. Poznáme:
- Chuťové receptory;
- Čuchové receptory.
- Fotoreceptory – patria sem sietnicové receptory, ktoré reagujú na svetlo určitých vlnových dĺžok.
- Nociceptory reagujú na pôsobenie nadmerne silných podnetov, ktoré poškodzujú tkanivá.
Senzorické dráhy
Všetky senzorické dráhy majú určité veci spoločné:
- Začínajú podnetom, ktorý vo forme fyzikálnej energie pôsobí na senzorický receptor;
- Receptor je transducer, ktorý konvertuje podnet na intracelulárny signál – akčný potenciál.
- Transdukcia je premena energie podnetu do informácie, ktorá môže byť spracovaná nervovým systémom. Ide v podstate o otváranie a zatváranie sa ionových kanálov citlivých na mechanickú, chemickú, tepelnú alebo svetelnú energiu.
- Ak je podnet prahový, akčný potenciál prechádza senzorickým neurónom do centrálneho nervového systému, kde je integrovaný.
- Senzorická informácia z tela vstupuje do spinálnej miechy a ide vzostupnými dráhami do mozgu. Niektoré senzorické informácie idú priamo do mozgu cez mozgový kmeň kraniálnymi nervami. Senzorická informácia, ktorá spúšťa viscerálne reflexy, je intergovaná v mozgovom kmeni alebo spinálnej mieche a obyčajne nedosahuje vedomé vnímanie.
Úloha CNS
CNS musí rozlíšiť 4 vlastnosti podnetu:
- Pôvod alebo modalitu – je indikovaná tým, ktoré senzorické neuróny sú aktivované a kde dráhy aktivovaných neurónov končia v mozgu. Každý receptorový typ je najcitlivejší na čiastočnú modalitu podnetu.
- Lokalizáciu – lokalizácia podnetu je kódovaná podľa recepčných polí, ktoré sú aktivované. Senzorické oblasti mozgu sú vysoko organizované s ohľadom na vstupné signály a vstup z priľahlých senzorických receptorov je spracovaný v priľahlých oblastiach kôry.
- Intenzitu – je určovaná jednak počtom aktivovaných receptorov ako aj frekvenciou akčných potenciálov prichádzajúcich z týchto receptorov
- Trvanie – je dané trvaním akčného potenciálu v senzorickom neuróne. Vo všeobecnosti platí, že dlhší podnet generuje dlhšiu sériu akčných potenciálov v primárnom senzorickom neuróne. Však ak podnet pretrváva, niektoré receptory sa adaptujú.
Zrak
Optický aparát oka je tvorený rohovkou, prednou komorou, šošovkou a sklovcom. Optická mohutnosť neakomodovaného oka je približne 59 D.
Optické princípy
- Zväzok rovnobežných lúčov sa spojuje v zadnom ohnisku;
- Lúč prechádzajúci predným ohniskom postupuje okom rovnobežne s optickou osou;
- Lúč identický s optickou osou sa neláme.
Mechanizmus akomodácie. Šošovka je pripevnená k musculus ciliaris neelastickými ligamentami. Keď je musculus ciliaris relaxovaný, ligamenta sú ťahané, vyrovnávajú sa a šošovka sa vyrovnáva. Akonáhle sa musculus ciliaris kontrahuje, uvoľňuje napätie ligament a šošovky sa zaokruhľujú.
Zraková ostrosť (visus) – je schopnosť rozoznať dva body v priestore s minimálnou vzájomnou vzdialenosťou a závisí na:
- Ich vzájomnej vzdialenosti,
- Vzdialenosti od očí, čo znamená od uhlu, ktorý je medzi lúčmi prichádzajúcimi od týchto bodov.
- Ak je tento uhol menší (minimum separabile), tak je zraková ostrosť väčšia.
- Hustote aktívnych fotoreceptorov.
Zrakové defekty
- Emetropické oko – normálne oko. Svetelné lúče dopadajúce do oka sa po prechode svetlolomnými plochami lámo do ohniska ležiaceho na sietnici.
- Myopické oko – krátkozraké oko. Defekt je spůsobený tým, že sa lúče lámu do ohniska, ktoré sa nachádza pred sietnicou. Chyba sa koriguje umiestnením rozptyliek pred oko.
- Hypermetrické oko – ďalekozraké oko. Defekt je spůsobený tým, že sa lúče lámu do ohniska za sietnicou. Chyba sa koriguje umiestnením spojky pred oko.
- Astigmatizmus – je spôsobený nerovnakým zakrivením očnej gule v horizontálnej a vertikálnej rovine.
- Škúlenie (strabizmus) – zaostrenie na blízky bod je sprostredkované konvergenciou očí a fixovaný objekt sa premieta do fovea centralis. Škúlenie pri hyperopii je nevyhnutné najmä pri pohľade do diaľky, kedy vizuálne osi konvergujú (u emetropického oka, vizuálne osi sú paralelné).
- Presbyopia – je zhoršená schopnosť zaostrenia (strata flexibility šošovky) u starších ľudí.
Sietnica (retina)
Na sietnici rozpoznávame:
- Macula lutea (centrum zrakového poľa);
- Fovea centralis (oblasť najostrejšieho videnia) – je tu najväčšia koncentrácia čapíkov;
- Optický disk (slepá škvrna) – miesto, kde sa nenachádzajú žiadne fotoreceptory. Je to miestom vstupu ciev a výstupu optického nervu.
Usporiadanie sietnice (obr. 2):

Obr. 2. Usporiadanie sietnice.
Receptory
- Čapíky – slúžia na videnie cez deň a na rozoznávanie farieb – fotopické videnie
- Tyčinky – slúžia na videnie za šera a umožňujú len neostré a bezfarebné videnie – skotopické videnie
Mechanizmus podráždenia
- V tme je rodopsín neaktívny, cGMP je vysoké a Na+ a K+ iónové kanále sú otvorené. Na+/K+- pumpa udržuje iónovú rovnováhu a membránový potenciál na -40 mV. Neurotransmiter sa uvoľňuje do bipolárnych buniek.
- Svetlo rodopsín rozštiepi, opsín znižuje cGMP, Na+ kanále sa zatvárajú, bunka sa hyperpolarizuje na -70 mV. Znižuje sa uvoľňovanie neurotransmiteru počas svetla.
- Vo fáze obnovy sa retinal rekombinuje s opsinom na inaktívnu formu rodopsinu.
Horizontálne bunky. Spájajú tyčinky a čapíky ako aj nasadajú na dendrity bipolárnych buniek. Výstup horizontálnych buniek je vždy inhibičný. Laterálne spojenie poskytuje rovnaký fenomén laterálnej inhibície, t.j. šíriaci sa excitačný signál v retine je horizontálnymi bunkami inhibovaný. Toto je významným mechanizmom, ktorý dovoľuje vizuálnu presnosť v prenose kontrastu hraníc sledovaného obrazu, ale pri veľkých vzdialenostiach.
Bipolárne bunky. Sú excitačné (depolarizujúce) a inhibičné (hyperpolarizujúce), čo dovoľuje jednej polovici bipolárnych buniek prenášať pozitívne a druhej negatívne signály, ktoré sú potom prenášané do mozgu. Iným významom tohoto reciprokého vzťahu medzi depolarizačnými a hyperpolarizačnými bipolárnymi bunkami je, že poskytujú sekundárny mechanizmus laterálnej inhibície. Kedže oba typy ležia oproti sebe navzájom, vytvára sa mechanizmus pre oddelený vizuálny kontrast aj keď hranica leží presne medzi dvoma priľahlými fotoreceptormi.
Existujú dva typy bipolárnych buniek:
- Light-on bipolárne bunky – počas tmy sú inhibované neurotransmiterom glutamátom, ktorý je uvoľňovaný z fotoreceptorov, počas svetla sú aktivované.
- Light-off bipolárne bunky – počas tmy sú excitované neurotransmiterom glutamátom, ktorý je uvoľňovaný z fotoreceptorov, počas svetla sú inhibované.
Či je glutamát excitačný alebo inhibičný závisí na type glutamátového receptora na bipolárnom neuróne.
Amakrinné bunky. Bolo určených asi 30 typov amakrinných buniek s rozličnými funkciami. Jeden typ amakrinných buniek je časťou priamej cesty od tyčiniek, cez bipolárne bunky a amakrinné bunky na gangliové bunky. Iný typ reaguje na začiatok vizuálneho signálu, ale reakcia odznieva rýchlo. Iné amakrinné bunky reagujú na vypnutie vizuálneho signálu a podobne reakcia odznieva pomerne rýchlo. Iný typ reaguje aj na prvú aj na druhú situáciu. Ďalší typ reaguje na pohyb svetelného bodu na sietnici v špecifickom smere – smerová citlivosť.
Gangliové bunky. Poznáme:
- Gangliové bunky s malým jadrom – P bunky – ktoré končia v štyroch dorzálnych vrstvách corpus geniculatum laterale. P systém prenáša najmä zrakové informácie o farbe, je málo citlivý na kontrast. Ale vyznačuje sa vysokou rozlišovacou schopnosťou.
- Gangliové bunky s veľkým jadrom – M bunky – končia v dvoch ventrálnych vrstvách corpus geniculatom laterale. M systém zabezpečuje je citlivejší na zmenu kontrastu a sprostredkúva najmä vnímanie pohybu predmetov v zornom poli.
Gangliové bunky majú spojenie s bipolárnymi bunkami, ktoré môžu byť excitačné alebo aj inhibičné, pričom každá gangliové bunka dostáva signál zo určitej oblasti sietnice – zrakové polia:
- Gangliové receptívne pole „on“. Ide o zapínacie gangliové bunky s excitačným centrom a inhibičnou perifériou.
- Gangliové receptívne pole „off“. Ide o vypínacie gangliové bunky s inhibičným centrom a excitačnou perifériou.
Zraková dráha
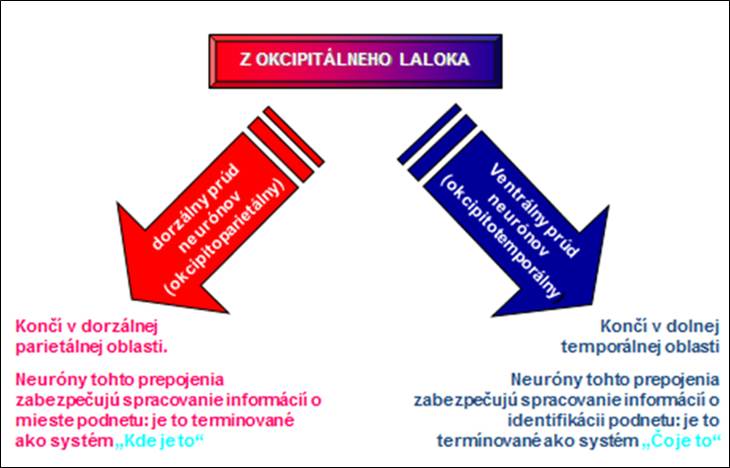
Vlákna n. opticus sa čiastočne krížia v chiasma opticum, pretože sa tu krížia len vlákna z nazálnych častí sietnice, ktoré prenášajú zrakové informácie z temporálnych častí zrakového poľa. Vlákna z temporálnych polovíc zorného poľa, prenášajúce informácie z nazálnych polovíc zorného poľa, sa nekrížia a pokračujú ipsilaterálne. To znamená, že v pravej zrakovej dráhe sú vlákna z temporálnej polovice sietnice pravého oka a nazálnej polovice sietnice ľavého oka a prenášajú sa ňou informácie z ľavej polovice poľa cez pravé corpus geniculatum laterale do pravej oblasti area 17.
Centrálny zrakový analyzátor na nachádza v sulcus calcarinus okcipitálneho laloka (area 17). V arei 18 a 19 dochádza k zrakovým asociáciam. Tieto oblasti majú eferentné spojenia s frontálnou oblasťou, ktorá má vzťah ku kontrole pohybov očných gúľ, reakciami dúhovky a slzením.
Obr. 3. Spracovanie zrakovej informácie.
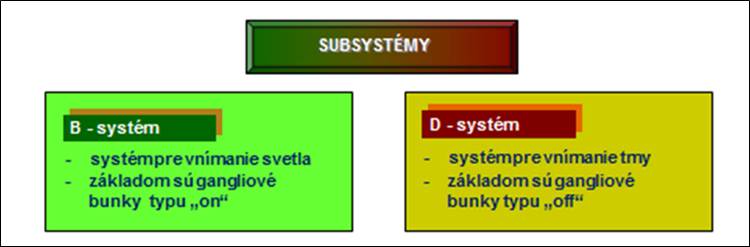
Obr. 4. Subsystémy spracovania zrakovej informácie
Vnímanie tmy je aktívny proces a nie len strata svetelného vnímania.
Farebné videnie
Farbocit – schopnosť správneho vnímania farieb okom.
Slnečné svetlo (biele svetlo) je polychromatická farba. Každá farba je daná farebným tónom, svietivosťou a sýtosťou.
Poznáme:
- Základné farby;
- Doplnkové farby.
Trichromatická teória – na sietnici máme 3 druhy čapíkov, citlivých na absorpčné maximá elektromagnetického vlnenia:
- V krátkovlnnej časti spektra – okolo 435 nm (cyanolab – pre modrú farbu)
- V strednej časti spektra – okolo 540 nm (chlorab – pre zelenú farbu)
- V dlhovlnnej časti spektra – okolo 570 nm (erytrolab – pre červenú farbu)
Pri podráždení len jedného druhu čapíkov máme vnem príslušnej základnej farby. Pri rovnomernom podráždení všetkých troch druhov čapíkov získaváme vnem bielej farby.
- Trichromacia – normálne farebné videnie.
- Dichromacia – normálne farebné videnie, prítomných je 2 druhy čapíkov.
- Monochromacia – prítomných je len 1 druh čapíkov, človek vníma len odtiene šedej farby, je vzácna, často komplikovaná svetloplachosťou, zníženou zrakovou ostrosťou a ďalšími poruchami zraku.
Poruchy farebného videnia
- Protanomalia – znížená citlivosť na červenú;
- Deuteroanomalia – znížená citlivosť na zelenú;
- Tritanomalia – znížená citlivosť na modrú;
- Protanopia – výpadok citlivosti na červenú;
- Deuteranopia – výpadok citlivosti na zelenú, mýlia si červenú a zelenú – vnímajú ich ako žltú;
- Tritanopia – výpadok citlivosti na modrú, nerozlišujú modrú a žltú.
Zrakové reflexy
Vôľové pohyby očí sú spúšťané v motorických centrách frontálneho laloka.
Reflexné pohyby očí sú spúšťané dvoma skupinami neurónov:
- Skupina reaguje na vizuálny podnet s centrom v okcipitálnom laloku.
- Skupina reaguje na nevizuálny podnet s centrom v strednom mozgu a ponte. Impulzy sa prenášajú III., IV. a VI. kraniálnym nervom.
Myotický (pupilokonstrikčný) zornicový reflex – parasympatikus - m. sphincter pupilae.
Mydriatický (pupilodilatačný) zornicový reflex – symapatikus – m. dilatator pupilae.
Sluch
Fyzikálne vlastnosti zvuku:
- Amplitúda – zodpovedá hlasitosti (intenzite) zvuku.
- Frekvencia – zodpovedá výške zvuku.
- Zastúpenie harmonických poprípade disharmonických zložiek – zodpovedá skladbe zvuku.
Prenos zvuku uchom:
- Prvý prenos - zvukové vlny narážajú na bubienok a spôsobujú vibrácie.
- Energia zvukových vĺn je prenášaná troma kostičkami stredného ucha, ktoré vibrujú.
- Druhý prenos - strmienok je pripojený na membránu oválneho okienka. Vibrácie oválneho okienka rozvlňujú tekutinu v kochlei.
- Tretí prenos - vlny tekutiny tlačia na flexibilné membrány kochlei. Vláskovité bunky sa ohýbajú a uvoľňujú neurotransmiter.
- Štvrtý prenos – neurotransmitery sa uvoľňujú do senzorických neurónov a vytvárajú akčné potenciály, ktoré idú kochleárnym nervom do mozgu.
- Energia vlnenia prechádza cez kochleárnu dutinu do tympanickej dutiny a rozptyľuje sa v strednom uchu na okrúhlom okienku.
Stredné ucho:
- Prevod zvuku – hydraulická transformácia (1 : 18).
- Ochranné reflexy – prah reflexov je asi 80 dB:
- M. stapedius - n. facialis - napínanie blany oválneho okienka;
- M. tensor tympani - n.trigeminus - napínanie bubienka.
Mechanizmus podráždenia
Receptormi sú vonkajšie a vnútorné vláskovité bunky nachádzajúce sa na bazilárnej membráne vo vnútornom uchu.
Ohnutie stereocílií na vláskovitých bunkách otvára apikálne K+ kanále, výsledkom čoho je depolarizácia. Depolarizácia umožňuje vstup Ca2+ iónov na bazálnom konci buniek. Vstup Ca2+ iónov spôsobje uvolnenie transmiteru a tak excitovať aferentné nervové vlákna.
Sluchová dráha
Zvukové vlny stimulujú receptorvé bunky v Cortiho orgáne ganglion spirale Corti → cochleárna vetva VIII. hlavového nervu → cochleárne jadrá (dorzálne a ventrálne) → nc. olivaris superior → lemniscus lateralis → coliculus inferior → corpus genitulatum mediale → sluchové centrum v temporálnom laloku.
Čuch (olfakcia, osmezis)
Funkcie čuchu:
- Príjem informácii z vonkajšieho prostredia;
- Ochranná funkcia – signál nebezpečie;
- Čchový – orientačný reflex;
- Vyhľadávanie potravy;
- Sekrécia tráviacich štiav – trávenie;
- Sexuálna funkcia – feromóny;
- Obranná – kýchanie a apnoe;
- - emócie;
- Signál chorôb;
- Aromaterapia.
Henningova klasifikácia pachov:
- Kvetový, ovocný, živicový, koreninový, hnilobný a spáleninový.
Súčasné systémy ale uvádzajú 50 typov senzácií a tiež toľko typov receptorov.
Adekvátnym podnetom sú látky rozpustené v hliene. Receptormi sú distančné chemoreceptory (vláskovité bunky) lokalizované v epiteli nosovej sliznice (5 cm2) a sú súčasťou primárneho senzorického neurónu.
Čuchová dráha
Aferentné vlákna ( axóny čuchových receptorov v n. olfactorius) tvoria terminály v bulbus olfactorius. Cez tractus olfactorius smerujú axóny do čuchovej kôry v uncus gyri parahippocampalis v temporálnom laloku. Čuchová dráha nemá prepojenie v thalame, ani čuchové centrum v neokortexe!!!! Dráha nejde cez thalamus.
Intenzita čuchovej senzácie zahŕňa chemickú charakteristiku látky, koncentráciu látky, rýchlosť prúdenia vzduchu a citlivosť receptorov (adaptácia).
Citlivosť receptorov stúpa s teplotou, je vyššia u žien, zvyšuje sa pri aktivácii sympatiku, je znížená u fajčiarov, zvyšuje sa čuchaním, zvyšuje sa pri inaktivácii iných zmyslov a vekom klesá.
Poruchy čuchu
- Hyposmia – zníženie čuchovej citlivosti;
- Anosmia – zánik čuchovej citlivosti;
- Hyperosmia – zvýšenie čuchovej citlivosti.
N. trigeminus – v nose prítomné zakončenia sú dráždené iritujúcimi látkami (chlór, mentol, čpavok), ktoré vyvolávajú reflexné reakcie – apnoe, kýchanie, slzenie.
Chuť (geúzia, gustácia)
Receptormi sú chemoreceptory, ktoré sú dráždené látkami rozpustenými v slinách. Sú v pohárikoch v papillae lingualis v sliznici jazyka, v sliznici epiglottis, podnebia a faryngu.
Chuťové podnety:
- Sladké – hrot jazyka;
- Slané – zadné okraje jazyka;
- Kyslé – predné okraje jazyka;
- Horké – koreň jazyka;
- Umami – bielkoviny mäsa, rýb, vajec, syrov;
- Doplnok – pálivé koreninové vnemy – spôsobujú miernu bolesť.
Chuťová dráha
Aferentné vlákna primárnych senzorických neurónov priliehajú na spodinu chuťovej bunky (50 vláken na 1 pohárik). Vlákna z predných 2/3 jazyka prebiehajú v chorda tympani a nervus facialis, zo zadnej strany v n. glossopharyngeus. Z receptorov mimo jazyka v n. vagus.
Všetky vlákna končia presynapticky v nucleus tractus solitarii v predĺženej mieche, axóny týchto sekundárnych buniek sa synapticky prepájajú na neuróny špecifických jadier thalamu, na tretí sériovo zapojený neurón. Ich axóny sa končia na projekčných kôrových neurónoch v gyrus postcentralis. Vlákna z ncl. tractus solitarii vedú aj k jadrám spúšťajúcim sekréciu slín (n. salivatorius superior et inferior).
Poruchy chute
- Hypogeuzia – pokles chuťovej aktivity;
- Ageuzia – zánik chuťovej aktivity;
- Hypergeuzia – zvýšenie chuťovej aktivity.
Somestetický analyzátor
Somestetická citlivosť predstavuje nervové mechanizmy zbierajúce senzorickú informáciu z tela.
Klasifikácia somestetickej citlivosti
- Bolestivá citlivosť, je aktivovaná mnohými faktormi, ktoré poškodzujú tkanivo.
- Mechanoreceptívna somestetická citlivosť, zahrňuje jednak taktilné a jednak pozičné vnímanie, receptory sú stimulované mechanickým podnetom.
- Termoreceptívna citlivosť, deteguje teplo a chlad.
Klasifikácia somestetických citlivostí z funkčného pohľadu
- Systém povrchovej kožnej citlivosti alebo komplexný exteroreceptívny analyzátor – dotyk, tlak, teplo, chlad a povrchová bolesť. Informuje nervový systém o všetkých zmenách pôsobiacich na povrch tela.
- Systém hlbokej citlivosti alebo komplexný proprioreceptívny analyzátor – svaly, šľachy, hlboká bolesť. Informuje nervový systém o momentálnom stave pohybového aparátu organizmu.

Obr. 5. Päť základních kožních citlivostí.
Dotyk a tlak
Receptory:
- Vater-Paciniho telieska – rýchla adaptácia, nedostatok smerovej orientácie, krátky cyklus zotavenia a nepatrná únava, možnosť reagovať na vibráciu.
- Golgi-Mazzoniho disky – podobné vlastnosti ako u Vater-Paciniho teliesok.
- Meissnerove telieska – rýchla adaptácia, bez smerovej orientácie.
- Receptory vlasových folikulov – pomalá adaptácia.
- Merkelove disky, dotykový disk alebo dotykový meniskus – pomalá adaptácia, reagujú na tlakový podnet a trvanie podnetu, smerová orientácia, vo vlasovej časti.
- Ruffiniho zakončenia – pomalá adaptácia, smerová orientácia.
Nervové vlákna pre dotykové a tlakové receptory:
- Myelinizované aferentné nervové vlákna, rýchlosť vedenia 30 – 72 m/s, patria do II. skupiny (Lloyd) alebo do skupiny A alfa (Gasser);
- Nemyelinizované aferentné nervové vlákna rýchlosť vedenia 0.4 – 0.2 m/s, patria do IV. skupiny (Lloyd) alebo do skupiny C (Gasser).
Dráhy pre hrubý dotyk a tlak z tela a končatín (protopatické čitie)
Dotykové a tlakové receptory v koži → spinálne ganglion (1. senzorický neurón) → zadné rohy (2. senzorický neurón) → vlákna prechádzajú strednú čiaru → ventrálny spinothalamický trakt → lemniscus nmeidalis → thalamus (3. senzorický neurón) → mozgová kôra gyrus postcentralis.
Dráhy pre jemný dotyk a tlak z tela a končatín (epikritické čitie)
Dotykové a tlakové receptory v koži → spinálne ganglion (1. senzorický neurón) → spinobulbárny trakt → ncl. cuneatus a gracilis (2. senzorický neurón) → lemniscus medialis → thalamus (3. senzorický neurón) → mozgová kôra gyrus postcentralis.
Propriocepcia
Propriocepcia je zvláštny zmysel. Bol objevený fyziológom C. S. Sherringtonom.
Receptormi sú svalové vretienka:
- Sú komplexným senzorickým receptorovým orgánom myotatického reflexu;
- Sú umiestnené vo vnútri kostrového svalu;
- Pozostávajú z dvoch špecifických druhov intrafuzálnych vlákien;
- Sú priložené paralelne k extrafuzálnym vláknam.
Intrafuzálne vlákna sú inervované γ-motoneurónmi, ktoré sa nachádzajú v ventrálnej časti šedej hmoty miechy.
Motorický γ-systém:
- Jeho hlavnou úlohou je regulácia citlivosti primárnych a sekundárnych zakončení svalového vretienka.
- Neuróny γ-systému fungujú ako odporový regulátor a pripravujú kostrový sval na pohyb.
Proprioceptívne dráhy
Receptory → spinálne ganglion (1. senzorický neurón) → ipsilaterálny spinobulbárny trakt → ncl. cuneatus (vlákna z hornej časti tela a končatín) a ncl. gracilis (vlákna z dolnej časti tela a končatín). → 2. senzorický neurón, ktorý prechádza na opačnú stranu mozgového kmeňa (lemniscus medialis) → thalamus (3. senzorický neurón) → mozgová kôra gyrus postcentralis a lobus parietalis, kde sa uvedomuje svalová a šľachová citlivosť.
Statická složka propriocepce
- Její hlavní část je vedena výše popsanou spinobulbární drahou z receptorů a spinálního ganglia (1. neuron; tzv. primární aferentní vlákna) přímo do stejnostranných (ipsilaterálních) nc. gracilis a nc. cuneatus.
- Druhá část je vedena vlákny ze spinálního ganglia, která jdou descendentně a ascendentně a přepojují se v Rexedově zóně IV zadních rohů (2. neuron). Odtud vystupuje axon a zanořuje se do zadních provazců. Směřuje s nimi k zona reticularis nc. gracilis anebo nc. cuneatus (tzv. sekundární aferentní vlákna), jež dále pokračují spolu s lemniscus medialis (3. neuron) do thalamu a pak (4. neuron) do kůry.
- Třetí významná část se vyčleňuje jako podmnožina sekundárních vláken – descendentní axony spinálního ganglia se větví v zadním míšním rohu a odtud po přepojení sekundární vlákno směřuje vzhůru a končí v nc. cervicalis lateralis (ve výši C1-C2); k tomuto jádru také směřují odbočky z tr. spinothalamicus anterolateralis – tj. signály bolesti, teple a protopatickém čití). Po přepojení (tj. 3 neuron) stoupá jako součást svazku tr. cervico-thalamicus spolu s lemniscus medialis do thalamu a odtud do kůry.
Kinetická složka propriocepce
- Z dolní končetiny se převážně přepojuje v nc. Stilling-Clark v zadních míšních rozích (2. neuron) a odtud vede v laterálních provazcích míchy jako kontralaterální tr. spinocerebellaris ventralis anebo jako ipsilaterální tr. spinocerebelaris dorsalis ascendentně do mozkového kmene a cestou brachia conjunctiva (ventrální dráha) anebo corpora restiformia (dorzální dráha) končí v mozečku.
- Kinestetické informace z dolní kočetiny, které vcházejí do našeho vědomí, jdou cestou přepojení v nc. „Z“ uloženého v medula oblongata dorzolaterálně od nc. gracilis, odkud pak s lemniscus medialis pokračují do thalamu a do kůry.
- Z horní končetiny postupuje kinestetická propriocepce cestou zadních provazců, přepojuje se v nc. cuneatus lateralis a odtud pokračuje jak do mozečku, tak do thalamu a do kůry velkého mozku.
Reflexy
Myotatický reflex je výhradne monosynaptický a uskutočňuje sa aktiváciou anulospirálnych zakončení, ktoré vysielajú impulzy cez aferentáciu Ia k α-motoneurónom homonymného svalu.
Inverzný myotatický reflex zabezpečuje plynulú koordináciu aktivity kostrového svalu a zabraňuje nadmernému natiahnutiu šliach. Receptormi sú Golgiho šľachové telieska, ktoré zabezpečujú deje podľa následujúcej schémy:
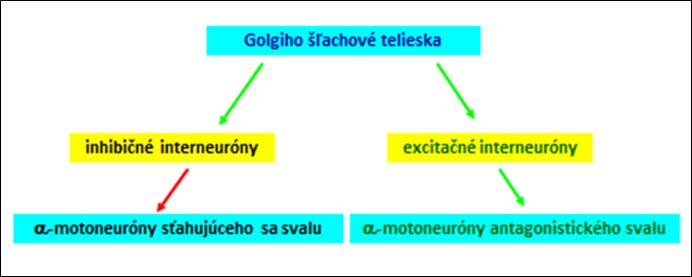
Obr. 6. Schéma činnosti zabezpečovanej Golgiho šľachovými telieskami.
Reciproký útlm
- Pri podráždení svalu agonistu aferentáciou Ia (myotatický reflex) dochádza súčasne k inhibícii anatagonistu;
- Uskutočňuje sa polysynaptickým vedením kolaterálmi Ia k alfa-motoneurónom antagonistického svalu.
Segmentálna regulácia
Jedná sa o mechanizmy spätnej väzby.
- Rekurentný útlm – k útlmu α-motoneurónov dochádza prostredníctvom skupiny inhibičných neurónov (Renshawove bunky), ktoré sú dráždené kolaterálmi týchto α-motoneurónov.
- Presynaptický útlm – jedná sa o útlm segmentových reflexných dráh. Dochádza k nemu, ak kolaterála aferentácie je v spojení s tlmivým interneurónom, ktorý môže znížiť výdaj transmiteru pre aferentáciu Ia k motoneurónom.
Supraspinálna regulácia
Podieľa sa významným spôsobom na regulácii svalového tonusu. Vyradenie tejto regulácie spôsobí v extenzoroch nadmernú reaktivitu myotatického reflexu a vedie k decerebračnej rigidite. S motoneurónmi a interneurónmi v mieche sú v spojení dráhy, ktoré majú hlavne dráždivé vplyvy na segmentovú reflexnú aktivitu, ale môžu pôsobiť aj tlmivo. Sú to reflexy:
- Supraspinálne
- Kortikospinálne
- Rubrospinálne
- Tektospinálne
- Olivospinálne
Rovnováha – vestibulárny aparát
- Vnímanie odchýlky postavenia hlavy vzhľadom na gravitáciu, pri zmene rýchlosti a smeru pohybu hlavy a celého organizmu v priestore, ako aj pri priamočiarom alebo kruhovom pohybe;
- Reflexná úprava tonusu najmä antigravitačných svalov, udržujúcich v vzpriamenej polohe hlavu a trup;
- Riadenie normálneho držania tela, rovnováhy tela a orientujú človeka o pohyboch organizmu v priestore a o zmenách parametrov týchto pohybov.
Vestibulárny aparát sa skladá:
- Z troch na seba kolmých, polkruhovitých kanálikov (canales semicirculares). Každý kanálik začína ampulou, do ktorej zasahuje hrana crista ampularis;
- Z dvoch váčkov – sacculus, utriculus. Obsahujú početné vápenaté telieska – otolity (macula utriculi, macula sacculi).
Statolitický aparát reaguje na zmenu polohy hlavy proti gravitácii:
- Sakulus – pri laterálnom úklone hlavy;
- Utrikulus – pri predklone a záklone hlavy.
Kinetický aparát reaguje na úhlové zrýchlenie, kde adekvátnym podnetom je zmena rýchlosti rotácie v rovine príslušného kanálika.
- Podráždenie vláskovitých buniek je výsledkom prúdenia endolymfy v kanáliku;
- Pohyb endolymfy ku crista amplularis vedie k depolarizácii vláskových buniek;
- Pohyb endolymfy od crista ampularis vedie k repolarizácii vláskových buniek.
Teplo a chlad
Receptormi sú termoreceptory, ktoré reagujú na zmeny teploty vo vonkajšom prostredí.
- Receptory na vnímanie tepla – voľné nervové zakončenie, generujú vzruchy v rozmedzí teplôt od 30 0C do 48 0C, nachádzajú sa v horných a stredných vrstvách choria.
- Receptory na vnímanie chladu – voľné nervové zakončenia, generujú vzruchy v rozmedzí teplôt od 10 0C do 45 0C, nachádzajú sa v epidermis kože.
Zvláštnosti termorecepcie:
- Reagujú len na relatívnu zmenu teploty okolia – indiferentná, fyziologická nulová teplota;
- Schopnosť generovať vzruchovú aktivitu pri konštantnej teplote okolia – statická reakcia;
- So zmenou teploty v čase sa ich vzruchová aktivita zvyšuje alebo tlmí – dynamický typ odpovede;
- Sú citlivé len na termické podnety.
Bolesť
Vnímanie bolesti spočíva v aktivácii zvláštnej skupiny nervových dráh, ktoré začínajú v periférnych nociceptoroch, teda v zmyslových orgánoch reagujúcich na škodlivé podnety. Premenlivosť reakcií na bolesť je daná veľkým počtom a zložitosťou ústredných kontrolných pochodov a značným počtom zúčastnených neurotransmiterov a neuromodulátorov.
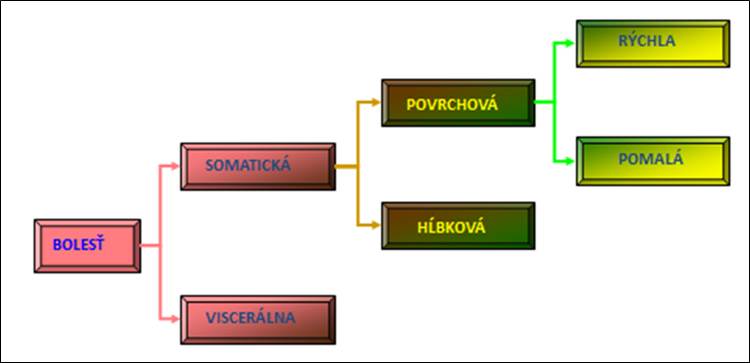
Obr. 7. Rozdelenie bolesti.
Receptory bolesti (nociceptory)
Receptory bolesti (nociceptory) sú voľné nervové zakončenia v koži, v peristole, stenách tepien, kĺboch a v tvrdej plene mozgu.
Poznáme:
- Mechanické nociceptory – receptory s A delta vlákna (15 – 25 m/s). Generujú vzruchy len pri pôsobení silných mechanických podnetov a aktivita závisí na intenzite poškodzujúceho podnetu. Chemická a termická stimulácia je bez efektu.
- Mechanicko – termické nociceptory
- Receptory s A-δ vláknami (4 – 10 m/s). Podnet – teplota viac ako 45 0C.
- Receptory s C vláknami. Podnet – teplota menej ako 15 0C a silný mechanický podnet.
- Polymodálne receptory s C vláknami. Podnet – silný mechanický podnet, chemický podnet a termický podnet od 42 0C do 60 0C.
Zmeny v pociťovaní bolesti
Naše vnímanie bolesti je objektom modulácie na niekoľkých úrovniach v nervovom systéme. Môže byť zväčšené poslednými skúsenosťami alebo potlačené nevyhnutnosťou, ak prežitie závisí na ignorovaní poškodenia. V poslednej situácii, zostupné dráhy prechádzajú cez thalamus a inhibujú nociceptorové neuróny v spinálnej mieche. Umelá stimulácia týchto inhibičných dráh je jedným z mála metód, ktoré sa používajú na reguláciu chronickej bolesti.
- Hypoalgézia – znížené vnímanie bolesti
- Hyperalgézia – zvýšené vnímanie bolesti
Tlumenie bolesti
Privieranie bolesti (pain gating) – nejde o zníženie citlivosti nociceptorov, ale o útlm prenosu senzorickej informácie do mozgu mechanizmom presynaptického útlmu. Keď sa súčasne s bolestivým dráždením nociceptorov dráždia aj dotykové alebo tlakové receptory, vnímanie bolesti sa oslabuje. Tento jav sa využíva pri intenzívnej bolesti – jemné masírovanie kože v blízkosti bolestivého miesta.
Mechanizmus:
- Bolesť môže byť potlačená v zadných rohoch miechy ešte pred prevedením impulzu vzostupnými spinálnymi dráhami. Normálne tonicky aktívne inhibičné interneuróny v spinálnej mieche inhibujú vzostupné dráhy pre bolesť. C vlákna majú synapsiu na inhibičných interneurónoch.
- Ak sa pôsobí bolestivým podnetom, C vlákna súčasne excitujú vzostupnú dráhu a blokujú tonickú inhibíciu, umožňujúc tak prenos signálu do mozgu.
- A-β vlákna prevádzajú senzorickú informáciu o mechanickom podnete a pomáhajú blokovať prenos bolesti. A-β vlákna majú synapsiu s inhibičnými interneurónmi a zosilňujú ich aktivitu. Ak súčasný podnet dosiahne inhibičný neurón z A-β a C vlákien, výsledkom je čiastočná inhibícia vzostupnej dráhy.
Centrálny analgetický systém (CAS)
Centrálny analgetický systém (CAS) sa môže aktivovať aj psychogénne (bolesť sa nepociťuje alebo sa pociťuje slabšie, ak poranenie nastane pri nejakej činnosti, ktorá si vyžaduje veľké sústredenie a motiváciu).
Centrálny analgetický systém tvoria tri hlavné a viaceré vedľajšie zložky (obr. 8):

Obr. 8. Zložky centrálneho analgetického systému.
Elektrická stimulácia štruktúr v prvých dvoch oblastiach môže úplne potlačiť signály bolesti vstupujúce do miechy.
Tieto štruktúry sa zvyčajne aktivujú z paraventrikulárnych jadier hypothalamu. V centrálnom analgetickom systéme sa ako transmitery uplatňujú enkefalíny (napr. v ncl. raphe magnus) a sérotonín, ktorý aktivuje neuróny v zadných rohoch miechy, kde sa opäť uvoľňuje enkefalín.
Enkefalíny (metenkefalín a leuenkefalín) tvoria spolu s β-endorfínom (v hypothalame a hypofýze) endogénny opiátový systém, ktorý má podobný analgetický účinok ako morfium a morfiové prípravky.
Vnímanie bolesti
Prah bolesti alebo jeho zmeny sú pociťované pochodmi v centrálnej aj periférnej nervovej sústave, pričom interakcia periférnych miechových a supraspinálnych systémov umožňuje organizmu využívať predchádzajúce skúsenosti ku kontrole senzitívnych vstupov na rôznych rovinách ústrednej nervovej sústavy (obr. 9, 10).
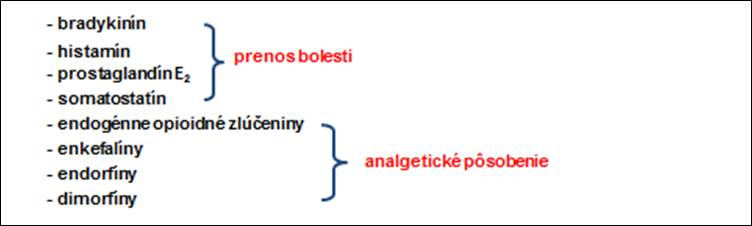
Obr. 9. Transmitery podieľajúce sa na nociceptívnych mechanizmoch.
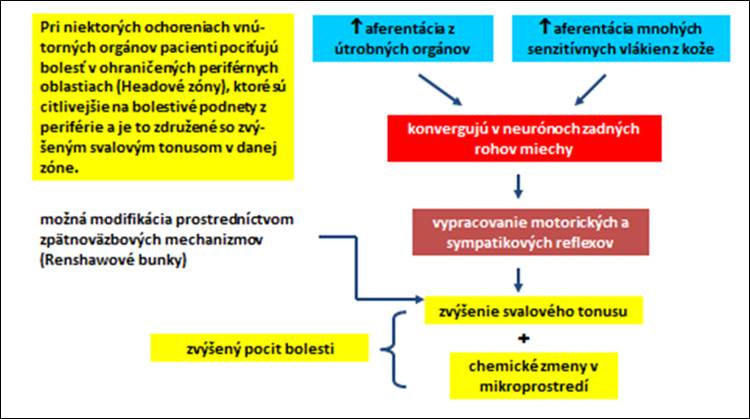
Obr. 10. Miechové mechanizmy nociceptorov.
Zvýšené svalové napätie je reakciou celého tela na bolesť a môže viesť, zvlášť v spinálnej oblasti, ku svalovému spazmu spôsobujúceho abnormálne ochranné držanie a tým i k ďalšej bolesti. Imobilizáciou príslušného spinálneho segmentu sa môže predísť vyvolaniu ďalšej bolesti.
Prenesená bolesť
Väčšinou ide o viscerálnu bolesť, ktorá sa nedá presne lokalizovať. Táto bolesť sa nepociťuje v časti tela, ktorá je jej zdrojom, ale na inom, často vzdialenom mieste. T
eória, ktorá to vysvetľuje (konvergenčná teória), vychádza z faktu, že vo všetkých periférnych nervoch je oveľa viac aferentných vlákien ako ascendentných vláken v miechových dráhach, ktoré prenášajú bolesť (najmä v anterolaterálnom trakte). Z toho vyplýva, že aferentné vlákna zo somatických štruktúr musia konvergovať tom istom miechovom neuróne ako aferentné vlákna z viscerálnych štruktúr. Bolesť zo somatických štruktúr je bežnejšia ako z viscerálnych orgánov a mozog identifikuje bolesť z viscerálneho orgánu tak akoby prichádzala zo somatickej štruktúry.
Dráha pre bolesť
Receptor v koži (1. senzorický neurón) → ganglion zadných koreňov spinálnej miechy → ganglion zadných rohov spinálnej miechy(2. senzorický neurón)(väčšina vlákien pak prechádza stredovou čiarou) → tr. spinothalamicus lateralis → cez lemniscus medialis v ponte → do thalamu (nižšie senzorické centrum) → (3. senzorický neurón) do somatosenzorickej kôry v gyrus postcentralis (vyššie senzorické centrum).
Spracoval: Doc. RNDr. Pavol Švorc, Ph.D., Katedra fyziologie a patofyziologie LF OU v Ostravě