Úvod
Otázka vědomí stále zůstává palčivým problémem soudobé vědy. Ať již jde o přírodní nebo humanitní vědy, patří otázka podstaty vědomí, jeho vzniku, smyslu a účelu k těm nejobtížnějším otázkám, na které není jednoduché nalézt uspokojivou odpověď. Tajemství mysli a vědomí bývá často srovnáváno s tajemstvím vesmíru jako celku. Přes přetrvávající a často nesmiřitelné diskuse na půdě vědy a filosofie o jednotlivých aspektech problému mysli a vědomí, převažují některé obecnější souvislosti, které jsou v rámci soudobého vědění (paradigmatu) preferovány. K těmto předpokladům patří evoluční původ vědomí. Vědomí se vyvinulo až u organismů, kteří byli potomky svých předchůdců, jež nebyli nositeli schopnosti vědomí. Vědomí je tak z hlediska evoluční biologie chápáno jako jeden z mnoha adaptačních znaků, který zvyšuje fitness jeho nositele a objevuje se v průběhu evolučního procesu jako adaptace až v pozdější době. Na druhé straně někteří evoluční biologové ale zpochybňují vznik vědomí jako přímou adaptaci a předpokládají, že mozek a vědomí spíše vznikly jako vedlejší efekt jiných změn (Gould a Lewontin 1979).
Evoluční původ vědomí
Přestože je evoluční původ vědomí obecně přijímaným faktem, je nicméně velice obtížné odpovědět na otázky, u kterých organismů se vědomí jako takové poprvé objevilo, jaké jim to přineslo evoluční výhody oproti jiným organismům, kteří touto schopností nedisponovali, a konečně ani to, jaké dědičné fyzické změny byly zodpovědné za vznik takové vlastnosti. Je tak obtížné zodpovědět otázku, jak se mohli již vyvinuté organismy bez cítění a vědomí vyvinout do organismů s cítěním a vědomím. Pravděpodobným předpokladem fyzických změn zodpovědných za vznik vědomé zkušenosti byly změny v anatomii a komplexitě mozku. Za primární formu vědomí je považováno percepční vědomí, které je východiskem pro vznik sebevědomí. Na rozdíl od toho, organismy s omezenými percepčními schopnostmi nemohou vytvářet vhodný model percepčního světa a vytvořit tak distinkci já a svět, která se zdá být základem vyšších forem vědomí a sebevědomí. Rozhodující pro vývoj vědomí jsou také společenské a kulturní souvislosti, role učení a jazyka.
Druhy vědomí
Není tedy pochyb o tom, že lidské vědomí je produktem evoluce jako postupného procesu, který nezačíná u přechodu od společného předka opice a člověka k člověku, ale pouze se v tomto přechodu završuje (de Duve 1995, 247). Lze tedy předpokládat, že evoluční vývoj vědomí sahá mnohem hlouběji do stále se rozvíjejícího evolučního stromu druhů a života a že může být přisouzeno i jiným druhům. Přestože pravděpodobně nemáme problém přisoudit jistý stupeň vědomí vyšším savcům, je otázka, kde stanovit hranici pro vědomé organismy a nevědomé organismy. Filosof Daniel Dennett zdůrazňuje, že tento problém má i morální a etické důsledky, protože pokud nepřiznáme vědomé percepční stavy některým životním formám, pak jim upíráme i stavy bolesti, což může ospravedlňovat neetické chování člověka vůči nim (Dennett [1997] 2004). Za rozumný tak lze pokládat předpoklad, že vědomí je stupňovitá a škálovitá vlastnost, kterou lze přisoudit mnoha živočišným formám a druhům od primitivních, elementárních a primárních vědomých funkcí až k plně rozvinutým schopnostem abstrakce, modelování, projekce a prožívání, jež používají sofistikované nástroje myšlení, především jazyka, učení a kulturního přenosu informací. Dennett vidí tento evoluční proces v několika základních krocích, pro které užívá analogii věže, jejíž nejspodnější patra obývají organismy s elementárními funkcemi vědomí až k patrům nejvyšším s vysoce rozvinutými sofistikovanými nástroji mysli (Dennett [1997] 2004).
Je-li evoluce, a tedy proces postupných modifikací předchozích znaků a vlastností organismů, zodpovědný za vznik vědomí jako pravděpodobně jednoho z nejsložitějších jevů a faktů našeho univerza, jakým způsobem probíhají tyto evoluční změny, které mají takový tvořivý potenciál, že jsou schopné vytvářet takto sofistikované vlastnosti a jevy? Jsou evoluční procesy jen záležitostí biologické říše nebo lze těchto procesů využít i v jiných oblastech skutečnosti? Pro možné odpovědi na takové otázky je třeba se podrobněji zabývat podstatou mechanismu evoluce a evolučních procesů jako takových a pokusit se charakterizovat jeho obecné vlastnosti a status v teoretickém myšlení. V této souvislosti nelze nepřihlédnoutk tomu, jakou roli sehrává evoluční koncept v kontextu soudobé vědy a filosofie.
Evoluce
Pojem evoluce je nepochybně jedním z centrálních pojmů soudobého myšlení, který podstatně ovlivňuje teoretické koncepce soudobé vědy a filosofie. Přestože se pojmu evoluce užívá v různých souvislostech a významech, jedná se původně o pojem „vývoje uzpůsobením“ (descent with modification), který vzešel z empirických objevů Charlese Darwina a Alfreda Russella Wallace. Tento původně darwinistický pojem evoluce v soudobé vědě funguje nejen jako základ všech biologických věd, ale objevuje se také v oblastech, které se netýkají výhradně života jako takového, a přesahuje tak oblast, v které byl původně objeven. Zdá se, že evoluce je přirozeným mechanismem, který má určitou univerzální podobu a plní tak explikační funkci v mnoha oblastech vědy.
Přírodní vědy
O této skutečnosti svědčí aplikovatelnost evolučního mechanismu nejen ve velice blízkých oborech biologie (např. genetice, morfologii, embryologii, geologii, ekologii, biogeografii, paleontologii), ale i ve vzdálenějších oborech, jako jsou chemie a fyzika (v hraničních oblastech biochemie a biofyzika), které se o pojem evoluce opírají a berou ho minimálně jako explicitně nevyjádřený předpoklad.
Počítačové vědy
V současné době je také nepochybné využití evolučního mechanismu v počítačových vědách, ať již v aplikační oblasti - při řešení takových problémů, které jsou obtížně řešitelné běžnými metodami, nebo v oblasti simulací, kde se testují vlastnosti evolučního mechanismu jako takového (genetické algoritmy, genetické programování, umělý život, digitální organismy).
Společenské vědy a filosofie
Také ve společenských vědách začíná evoluce hrát důležitou roli prostřednictvím úvah o formování kultury v rámci sociobiologie a nověji evoluční psychologie. V tomto kontextu se objevují výzkumy procesů jak kulturního přenosu, tak i adaptivity kultury. I v oblasti filosofického myšlení je pozoruhodný příklon v poslední době k naturalismu, a konkrétně evolučnímu naturalismu (Lorenz, Campbell, Toulmin, Popper, Quine, Kornblith, Laudan, a mnozí další). Evoluční epistemologie v současné době reprezentuje hlavní proud filosofického studia poznání a podobně ústřední roli hrají také evoluční mechanismy v teoriích vědomí, mysli, učení a morálky.
Role evoluce v soudobé vědě: explanace, unifikace, univerzalizace
Pojem evoluce splňuje v soudobé vědě základní funkce podobně jako jiné obecné termíny vědy.
- Především vystupuje jako univerzální explanační nástroj, jehož prostřednictvím lze vysvětlit nejen bohatství jevových biologických forem, jejich neobvyklou mnohotvárnost a zdánlivě „promyšlenou“ závislost a zřetězenost, ale také mnoho dalších procesů mimo výhradně biologickou oblast živého, ať již jde o oblasti přírodních nebo společenských věd.
- V této souvislosti je nutné si uvědomit univerzálnost takového evolučního mechanismu, jenž nastává vždy a všude, kde jsou k tomu stanoveny vhodné podmínky. Univerzalita takového procesu pak vede ke snaze formulovat takový proces nezávisle na jeho biologické instanci, a pochopit tak roli biologické evoluce jako jednoho z možných projevů mnohem obecnějšího přírodního procesu. Obecnost pak umožňuje formulovat predikce o možných, dosud nepoznaných příčinách nebo výsledcích procesů nebo o oblastech, kde lze očekávat jeho uskutečnění.
- Pojem evoluce konečně také sehrává roli unifikačního nástroje vědy, jenž sjednocuje nebývalou rozmanitost přírodních jevů v různých oblastech skutečnosti na základě jednoduchého a opakujícího se vzoru anbo zákona. V tomto smyslu může být evoluce porovnána s obdobnými unifikačními funkcemi jiných pojmů přírodních věd, například s gravitací, díky níž se Newtonovi podařilo sjednotit rozmanitost pohybů od oběhů planet a měsíců až k šikmému vrhu a pádu těles.
Co je evoluce?
Přestože je evoluční koncepce široce přijímána jako empirický a teoretický fakt anebo alespoň jako implicitní předpoklad, neexistuje konsensus o tom, co je evoluce nebo evoluční princip. V rámci diskusí v soudobé vědě se objevují dokonce vážné pochybnosti o darwinovském evolučním mechanismu – o roli přírodního výběru, o četnosti a významu adaptací, o evoluční zdatnosti atd. Lze však říci, že přestože jsou často tyto základní prvky darwinovského evolučního mechanismu zpochybňovány v kontextu nových objevů (genetika, teorie přerušovaných rovnováh, Evo-Devo) dochází později k syntéze s původním darwinismem. I když tedy výzkum směřuje k postupnému odhalování hlubších mechanismů, které Darwin nemohl znát (genetika, Evo-Devo), či k dalším aspektům evolučního mechanismu, není v zásadě prokázáno nic, co by bylo v přímém rozporu s darwinismem. Obecné přijetí darwinismu jako podstaty evoluce je tedy faktem soudobé vědy. Na druhé straně je ale pravdou, že přestože je evoluční teorie obecně přijímána, je dokonce i v rámci vědecké komunity skutečně pochopena jen výjimečně. Filosof biologie Robert Brandon považuje za zdroj tohoto problému sám pojem evoluce (Brandon 1978, 96).
Evoluci lze například definovat tak, že jde o „změnu v genotypu populace“ (Dobzhansky 1937, cit. podle Brandon 1978, 96). Brandon se domnívá, že je to velice vágní definice, která připouští rozmanité, a dokonce i paradoxní interpretace. Jedná se především o to, že jako změna v genotypu může být bráno cokoli včetně zrození i smrti individua. Zpřesněním a explicitním vyjádřením toho, co měl pravděpodobně původně Dobzhansky na mysli, dostáváme stávající a převažující definici evoluce: „Evoluce je jakákoli změna v relativní četnosti genů v populaci během času.“ (Brandon 1978, 96).
Podobně může být podstata biologické evoluce vyjádřena následujícími způsoby: Například: „Evoluce může být přesně definována jako jakákoli změna v četnosti alel v genetickém fondu z generace na generaci.“ (Curtis a Barnes 1989, 974). Nebo podobně: „Změny v populacích, které jsou brány za evoluční, jsou ty, které mohou být prostřednictvím genů děděny z jedné generace na druhou.“ (Futuyma 2005, 2).
V tomto typu definicí převažuje snaha určit evoluci prostřednictvím změn na genové úrovni (mikroúroveň), která je brána jako rozhodující pro změny ve fenotypu organismů (makroúroveň). V jistém smyslu je tento typ definice důsledkem odklonu od původní perspektivy, kdy za jednotku evoluční změny byl považován organismus, případně populace organismů v rámci druhu, a příklonu k tzv. „genocentrismu“, kdy díky objevu dědičného mechanismu byla upřena pozornost na geny jako na základní jednotky evoluce. Z hlediska současného biologického paradigmatu je to pochopitelné, neboť skutečně to jediné, co se zachovává, jsou alely jako konkrétní formy genů. Je to však důvodem i pro odpovídající definici evoluce?
Samozřejmě ne všechny definice evoluce důsledně respektují tuto moderní perspektivu genocentrismu. Např. v respektované Evoluci M. Ridleyho se říká:
- „Evoluce znamená změnu, změnu ve formě a chování organismů mezi generacemi. Formy organismů na všech úrovních od DNA sekvencí k makroskopické morfologii a sociálnímu chování mohou být modifikovány z jejich předchůdců během evoluce, ačkoli ne všechny typy biologických změn jsou zahrnuty do definice. Vývojová změna během života organismu není evolucí ve striktním smyslu […], změna ve složení počtu druhů v ekosystému by také nebyla běžně brána jako evoluce. […] Změny mezi generacemi v populacích druhů […] budeme nazývat evolucí. “ (Ridley 2004, 4; kurzíva v originále).
Kontextuální pojetí a evoluční definice
Je důležité uvědomit si, že i snahy o vymezení a definování evoluce jsou poplatné přetrvávající nejednotnosti v základních otázkách biologie. Tato názorová ideodiverzita (věrně kopírující výsledky biologické evoluce – biodiverzity – na jiné úrovni) je důsledkem specifičnosti a kontextuálnosti biologických jevů. Specifičnost vyplývá z autonomního postavení biologie jako přírodní vědy, již nelze jednoduše redukovat na chemické a fyzikální procesy, kontextuálnost potom vyplývá z rozmanitosti perspektiv, jež je možné užít pro popis a vysvětlení takto specifických dějů a procesů.
Multiúrovňovitost
V uvažovaném případě definování evoluce se tak nabízejí různé kontexty, v kterých může být podstata evolučního procesu vyjádřena. Z hlediska multiúrovňovitosti bychom mohli hovořit o mikroúrovňovém kontextu genetických změn, jež se dědí v dané linii generací, o makroúrovňovém kontextu morfologie a chování organismů v rámci populací a druhů, a případně i o výšeúrovňových kontextech úspěšných, či neúspěšných adaptací celých druhů i vyšších taxonů dané systematizace. Tento příklad je jen ilustrací dalších možných kontextuálních perspektiv, které je možné v rámci popisu a vysvětlení biologických dějů přijmout. Je z něho také patrné, že ne všechny kontexty, které mohou být uvedeny, jsou stejně významné a dosahují stejné hodnoty.
Z hlediska kontextuálního pojetí je tak třeba vnímat i pokusy o jednotlivé definice evoluce. Je evidentní, že zmíněné typy definic jsou buď příliš úzké, nebo naopak široké, a že všechny musí čelit opodstatněným námitkám. To však neznamená, že nesmiřitelnost mikroúrovňových a makroúrovňových pojetí je nepřekonatelná. Naopak z hlediska kontextuality je tato rozmanitost předpokládána, aniž by vedla k rozpadu a roztříštění jednotného obrazu světa na nesouvisející fragmenty. Rozmanitost neznemožňuje, ale naopak umožňuje vytváření souvislostí, jež vypovídají o charakteru sledované skutečnosti mnohem úplněji než jednostranné preferování toho či onoho kontextu na straně jedné nebo naopak pluralizace možných vysvětlení zamlčením vzájemných vazeb mezi nimi a přenesení explanační schopnosti na všechny kontexty rovným dílem na straně druhé. Pro snahy o nalezení definice evoluce to znamená, že je třeba nejen respektovat jak mikroúrovňové, tak makroúrovňové kontexty, ale že je také třeba soustředit se na strukturu vazeb, v kterých se tyto kontexty nacházejí a v některých případech dokonce předpokládají. Je tak pochopitelné, že makroskopický kontext, definující evoluci jako změnu ve formě a chování organismů mezi generacemi, nutně předpokládá mikroskopický kontext, definující tyto změny prostřednictvím kauzálních mechanismů na nižší úrovni, například jako změnu v relativní četnosti genů v populaci během času.
Jinými slovy, v době, kdy Darwin definoval evoluci z hlediska přírodního výběru a nebyl znám mechanismus dědičnosti prostřednictvím genů, bylo možné definovat evoluční změny probíhající na fenomenologické úrovni té doby (změnami forem a chování organismů v populacích organismů), aniž by to bylo v rozporu s možností definovat tyto změny prostřednictvím kauzálních mechanismů na nižší (genetické) úrovni. Je dokonce možné říci, že makroúrovňová definice evoluce předpokládá nutné kauzální síly na nižší než fenotypové úrovni, jež dědičnost jednotlivých znaků organismů zdůvodňují, a že tedy mikroúrovňový kontext je předpokladem makroúrovňového kontextu. Je však nutné poznamenat, že vzájemná podmíněnost těchto kontextů není symetrická. Mikroúrovňový kontext relativní četnosti genů v populacích nepředpokládá nutně existenci makroúrovňového kontextu fenotypové proměnlivosti organismů.
Evoluční a vývojové změny
Podstatným rysem biologické evoluce je to, že se netýká proměn (vývoje) individuálního organismu. Ontogenetické změny organismu jsou realizací genetického programu, který se předává z generace na generaci s více nebo méně významnými odchylkami. Původní Haeckelova představa, že ontogeneze (vývoj individuálního jedince) rekapituluje fylogenezi (vývoj druhu), je dnes již odmítnuta. Mezi ontogenezí a fylogenezí však existují vazby, které je nutné vzít v úvahu při specifikaci evolučních změn. Evoluční změny se sice projevují na každém individuálním jedinci (organismu) vzhledem k jeho předchůdcům a následníkům, ale o evoluční změny jde právě jen v časově rozsáhlejších obdobích, než je životní cyklus od zrození ke smrti individuálního organismu.
- „Biologická evoluce […] je změna ve vlastnostech populací organismů, která překračuje život jednotlivého individua. Ontogeneze individua není evolucí; individuální organismus se evolučně neproměňuje. Evoluční změny v populacích jsou ty, které jsou dědičné geneticky z jedné generace na další. Biologická evoluce může být nepatrná nebo podstatná; zahrnuje vše od nepatrných změn v proporcích různých alel uvnitř populace (např. těch, které určují krevní skupinu) až k následným změnám, které vedou od prvních protoorganismů k plžům, včelám, žirafám a pampeliškám.“ (Futuyma 1986, 7).
Nepředpokládáme tedy, že by bylo možné podat nějakou přesnější definici biologické evoluce mimo samotnou biologii. Zpřesnění takové definice je jistě záležitostí současného biologického paradigmatu nebo současné biologické racionality. Existuje však možnost pokusit se o charakteristiku základních principů, jež vedou k evolučnímu procesu, a ukázat také obecnější než jen čistě biologickou perspektivu evoluce jako přírodního procesu.
Nebiologická perspektiva evoluce
Ve skutečnosti se totiž problém evoluce neomezuje pouze na oblast biologických jevů. „V nejširším smyslu je evoluce pouze vše prostupující změna; galaxie, jazyky a politické systémy, vše evolvuje.“ (Futuyma 1986, 7). Je tak otázka, zda snaha definovat evoluci prostřednictvím specificky biologických souvislostí (genotyp – fenotyp, organismus – druh, adaptace – speciace apod.) není poněkud zavádějící - zda takové definice nejsou až příliš ovlivněny úzce vyhraněnou specifičností evoluce biologických druhů. Pro takový názor existuje několik důvodů:
- V rámci biologie existují fundamentální obtíže s definováním konceptu biologických druhů, přičemž selhává většina strategií, jak zdůvodnit původní linnéovskou taxonomii. Díky tomu pak evoluce jako změna probíhající v prostředí (populací) biologických druhů ztrácí oporu s tím, jak se pojem biologického druhu stává nejasným. Řešením takových obtíží pak je zdůraznění genetického základu dědičnosti a snaha definovat evoluci pouze prostřednictvím změn v četnosti genů v populaci.
- Ukazuje se, že v rámci biologie probíhají evoluční procesy, které se podstatně odlišují od evolučního procesu, jenž počítá s biologickými druhy – např. evoluce virů probíhá podle některých předpokladů vzhledem ke specifičnosti tzv. HGT (horizontálního genového transferu) bez formování druhů. I tyto procesy však podléhají přírodnímu výběru a genetickému základu informace a dědičnosti a formují v čase evoluční změny. Je tedy třeba uvažovat o evoluci i v kontextu těchto procesů a neomezovat se pouze na vyhraněnou část biologické skutečnosti, představovanou sexuální reprodukcí, jež umožňuje jen jeden z několika typů přenosu genetické informace.
- Konečně jsou také evoluční změny běžně přisuzovány široké třídě jevů od evoluce vesmíru až po evoluci kultury a je legitimní otázkou, zda se tento typ změn nějak podstatně liší od vyhraněně biologických procesů, nebo zda je možné nahlédnout tyto procesy v nějaké obecnější unifikující perspektivě.
Generalizace a axiomatizace evolučního procesu
Snaha strukturu a základní principy evolučního procesu nějak generalizovat pochází z přirozeného vědeckého úsilí o unifikaci rozmanitostí světa a jejího vyjádření prostřednictvím obecných a nutných vzorů, tj. zákonů. I v případě Darwinovy podoby evoluční teorie existují více či méně exaktnější pokusy o rekonstrukci fundamentálních principů teorie, jež by vedly k deduktivním důsledkům evolučních procesů. Podle některých interpretací Darwinova díla jsou základní čtyři principy uvedeny a obhájeny na počátku díla O původu druhů a pak je z nich dále odvozen princip přírodního výběru jako jejich důsledek. Tyto interpretace vidí Darwinovu teorii jako příklad klasického modelu deduktivní teorie, kdy ze základních principů (axiomů) odvozujeme známé i neznámé důsledky a další teorémy dané teorie. Takovou strukturu vykazuje nejen známá Eukleidova teorie geometrie a Newtonova mechanika, ale i mnoho dalších, především matematických a fyzikálních teorií.
Některé pokusy o generalizaci základních principů evolučního procesu byly již učiněny. Jedná se především o pokus o axiomatizaci evoluce (Mary B. Williams), reflexi obecných principů evoluce (Philip Kitcher), koncepci obecné evoluční změny (Donald Campbell), koncepci univerzálního darwinismu (Richard Dawkins, Daniel Dennett, Henry Plotkin) a ideu obecné formy selekčních procesů (David Hull).
Darwinova evoluční teorie jako východisko
Východiskem pro charakteristiku evoluce jako univerzálního přírodního procesu je formulace základních principů obsažených v díle Charlese Darwina O vzniku druhů přírodním výběrem z roku 1859. Ernst Mayr rozlišuje v původní Darwinově teorii pět principů nebo teorií, z nichž přijetí prvních dvou, tj. proměnlivosti druhů a vzniku ze společného předka, považuje za první darwinovskou revoluci, k níž došlo brzy po vydání jeho díla, zatímco přijetí dalších třech principů, tj. gradualismu, populačního vzniku druhů a přírodního výběru, spojuje až s druhou darwinovskou revolucí v evoluční syntéze. Je také třeba mít v této souvislosti na paměti Darwinovo původní odmítnutí Lamarckovského mechanismu dědičnosti získaných znaků a tzv. neodarwinismus Weismanna a Wallaceho, kteří ho popírali také. Z těchto důvodů Mayr dává přednost nazývat novodobý darwinismus, který se ujal po evoluční syntéze jednoduše darwinismem, protože je v klíčových aspektech v souladu s původním Darwinovým pojetím z roku 1859.
Vědecký status Darwinovy teorie
Darwinovo dílo O vzniku druhů přírodním výběrem, které je obecně přijímané jako Darwinova formulace teorie přírodního výběru, se také stalo objektem mnoha analýz ve filosofii vědy. Záměrem těchto analýz bylo a je zjistit, zda Darwinova teorie splňuje požadavky, které jsou kladeny na teoretický systém, který má být vědecký, a případně jakému modelu takového teoretického systému Darwinova teorie odpovídá. V této souvislosti lze říci, že se filosofové pokusili aplikovat všechny známé prostředky k tomu, aby se pokusili vyjasnit logickou strukturu Darwinova argumentu obsaženého v jeho díle a tím tak prokázat nejen vědeckost této teorie, ale také její induktivní nebo deduktivní strukturu.
Např. Karl Popper se nejprve domníval, že Darwinova teorie nesplňuje jeho falsifikační kritéria vědeckosti a přisoudil jí pouhý status metafyzického výzkumného programu. Tento svůj omyl ale brzy odvolal a podstatně revidoval své původní přesvědčení o netestovatelnosti Darwinovy evoluční teorie a jejím tautologickém charakteru. Změnil tak názor na logický status teorie přírodního výběru. Sám vždy považoval vědecký zájem o tuto teorii za oprávněný, nejen pro její enormní vysvětlující sílu, ale také proto, že z nejobecnějšího hlediska se mu otázka statusu vědeckosti Darwinovy teorie ukazuje jako teorie pokusu a eliminace chyb, tedy jeho základní postup při dosahování pravdy (Popper 1974, 195). Kromě Popperových názorů pak existuje celé spektrum hodnotících koncepcí od tradičního hypoteticko-deduktivního modelu teorie (Ruse 1975, Ghiselin 1984), přes model inference k nejlepšímu vysvětlení (Thagard 1978), model pravděpodobnostní kauzální teorie (Hodge 1977, 1987) a teorie sil (Sober 1984) až k sémantickému modelu teorie (Lloyd 1983, Sintonen 1990), k vysvětlující unifikaci (Kitcher 1985), logické struktuře kuhnovského paradigmatu (Wilson 1992) anebo k mechanistickému vysvětlení založeném na pojetí přírodního výběru jako mechanismu (Barros 2008).
Deduktivní charakter Darwinovy teorie
Jedním z těchto aspektů analýz Darwinovy teorie je předpoklad, že Darwinova teorie jako jádro darwinismu je buď plně, nebo alespoň částečně axiomatizovatelná, a že tak odpovídá tradičnímu hypoteticko-deduktivnímu modelu vědeckých teorií. Problémem pro taková pojetí je, že ne všechny části Původu druhů se dají jednoduše axiomaticky reformulovat tak, aby splňovaly deduktivní strukturu vědecké teorie. Jedná se především o ta místa, kde Darwin argumentuje na základě analogie mezi umělou a přirozenou selekcí, protože odpovídají spíše induktivní metodě úvahy. Některé autory to vede k závěru, že je to z hlediska hypoteticko-deduktivního ideálu vědy závažný nedostatek, který snižuje váhu a dopad Darwinovy teorie. Lze však ukázat, že induktivní části Původu druhů jsou více méně pragmatickou součástí Darwinova záměru oslovit čtenáře a nijak nesnižují deduktivní strukturu jeho teorie (Havlík 2011).
Jedním z filosofů, který v diskusích o vědeckosti Darwinovy teorie hájí její axiomatizovatelné jádro, je Philip Kitcher. Tvrdí, že ať již jsou diskuse o Darwinově teorii z hlediska statusu její vědeckosti jakkoli rozmanité a skeptické (Havlík 2008), většina musí souhlasit, že „každá vědecká teorie hodna toho jména musí obsahovat množinu tvrzení, mezi kterými jsou nějaké obecné zákony (zákony, které vyjasňují nejfundamentálnější regulativy ve zkoumané oblasti přírodních jevů), a že takové zákony musí být použity k odvození dříve nepřijatelných tvrzení, jejichž pravdivostní hodnota je předmětem empirického určení“ (Kitcher 1985, 129). V Darwinově případě pak předpokládáme, říká Kitcher, že „původ [druhů] předkládá nějakou kolekci nových obecných principů o organismech“ (Kitcher 1985, 129). Darwinova teorie tak spočívá na následujících čtyřech fundamentálních tvrzeních:
- V každé fázi historie druhu se budou nacházet variace mezi jeho členy; odlišné organismy patřící k druhu budou mít odlišné vlastnosti (Princip variace).
- V každé fázi historie druhu se rodí více organismů, než může přežít a reprodukovat se (Princip úsilí o existenci).
- V každé fázi historie druhu jsou nějaké variace mezi jeho členy, jež mají vztah k vlastnostem, které ovlivňují schopnost přežít a reprodukovat se; některé organismy mají charakteristiky, které je lépe připravují k přežití a reprodukci (Princip variace zdatnosti – in fitness).
- Dědičnost je pravidlem; většina vlastností organismů je děděna jejich potomky (Přísný princip dědičnosti) (Kitcher 1985, 129).
Podle Kitchera je možné rekonstruovat Darwinovu teorii na základě jeho díla O původu druhů tímto způsobem: první čtyři výše uvedené principy jsou obhájeny na počátku díla a pak je z nich odvozen princip přírodního výběru jako jejich důsledku. Kitcher se pro oprávněnost takové rekonstrukce odvolává také na řadu interpretů od T. H. Huxleyho až po Richarda Lewontina, kteří rekonstruují „jádro“ Darwinovy teorie stejným způsobem.
Princip přírodního výběru
I přes jeho notorickou známost zopakujme ve stručnosti tento přirozený neboli přírodní mechanismus evoluce. K výchozím faktům a empirickým evidencím, které měl na základě podrobného pozorování a studia Darwin k dispozici, patřily existence rozmanitosti druhů, jejich proměnlivost (variabilita) a schopnost přenášet určité znaky na potomky (dědičnost). Důkazy variability Darwin opíral o domestikaci zvířat a kultivaci rostlin, kdy se prokazovalo nejen to, že dědičné modifikace jsou možné, ale zároveň i to, že výběrem lze akumulovat následné jemné variace. Logika však musela být schopna podat nejen vysvětlení zrodu dokonalé formy druhu, ale především musela vysvětlit otázku koadaptace organismů navzájem a vůči prostředí jejich života. Zde si Darwin jasně uvědomoval komplexitu a složitost těchto vazeb a závislostí. Rozhodujícím impulsem mu pak byla Malthusova doktrína aplikovaná na celou živočišnou a rostlinnou říši, předpokládající geometrický růst organismů v omezeném prostředí s omezenými zdroji, jehož důsledkem musí být usilování o existenci. „Z toho, že se rodí mnohem více jedinců, než může přežít, a čehož důsledkem je neustálé usilování o existenci, vyplývá, že každá bytost, i když se liší jen drobně v jakémkoli způsobu, který ji může být prospěšný v celkových a měnících se podmínkách života, bude mít lepší šanci na přežití, a tedy bude přirozeně vybrána.“ (Darwin 1859).
Deduktivní struktura Darwinova argumentu
Obdobně jako v Kitcherově rekonstrukci Darwinova díla lze uvažovat možnou logicky-deduktivní strukturu úvahy, která tvoří jádro darwinismu. K výchozím axiomům tedy patří empirické evidence
- proměnlivosti druhů,
- dědičnosti znaků, z nichž alespoň některé mají vliv na přežití a schopnost další reprodukce,
- reprodukce organismů geometrickým růstem,
- konečný prostor s omezenými zdroji.
Logicko-deduktivním důsledkem těchto premis anebo axiomů pak je fakt evoluce přírodním výběrem. V tomto smyslu je evoluční proces zákonitým důsledkem, který nastává nutně a vždy, jsou-li splněny dané předpoklady. Teoretická axiomatizovaná struktura evolučního procesu je tak zobecněna, unifikována a abstrahována obdobným způsobem jako při formulaci např. fyzikálního zákona gravitace. Tento postup kromě jiného umožňuje i formulaci tzv. univerzálního darwinismu neboli univerzální evoluční perspektivy (Havlík 2008). Evoluce přírodním výběrem se na základě platnosti těchto univerzálních podmínek stává přírodním zákonem.
Univerzální darwinismus
Zmiňovaná axiomatizace je však jen jedním z mnoha pokusů o promyšlení univerzálních souvislostí evolučních mechanismů. Další takovou konceptuální možností je unifikující perspektiva „univerzálního darwinismu“ (Dawkins [1989] 2003; 1983; [1986] 2002), která rozšiřuje platnost Darwinovy evoluční teorie mimo hranice nám známé biologické evoluce. Dawkins není prvním, kdo myšlenku evoluce jako univerzálního přírodního zákona hájí. Sám se hlásí k tomu, že navazuje na myšlenky Poppera, Cavalli-Sforzy, Cloaka a Cullena, kteří poukázali na analogii mezi kulturní a genetickou evolucí (Dawkins [1989] 2003, 173). Dawkins ale nemá na mysli pouze tuto (pro mnohé často kontroverzní) analogii mezi biologickým a kulturním, ale mnohem univerzálnější pojetí evolučního procesu. Již v Sobeckém genu si klade otázku, zda existuje nějaký obecný princip, který platí pro veškerý život, zda existují pro život nějaké biologické principy s univerzální platností, ať již je tato forma života jakákoli, tedy i hypoteticky mimozemská. Dawkins tak klade otázku, zda můžeme chápat evoluci jako každý jiný rozumný přírodní zákon, jehož realizace je v rámci platnosti daných podmínek nutností.
Jestliže je tato Darwinovská rekonstrukce odvozena z konkrétní instance evolučního procesu, jehož jsme svědky zde na Zemi, pak je tu legitimní otázka po hranicích platnosti této Darwinem objevené zákonitosti. Dawkins je tím, kdo tuto otázku nejen artikuluje, ale také tím, kdo je ochoten vsadit na jeden základní princip, „na zákon, že každý život se musí vyvíjet na základě rozdílného přežívání replikujících se entit“ (Dawkins [1989] 2003, 174). (Pozn.: V českém překladu z roku 2003 je užito obratu „množících se věcí“, což vzhledem ke kontextu není úplně šťastné.) Dawkins modifikací „reprodukujících se organismů“ na „replikující se entity“ získává požadovanou univerzalizaci zákonitosti evolučního procesu. Replikující se entitou tak již není ani biologický organismus, ani jen živá entita jakékoli formy života ve vesmíru, ale skutečně libovolná entita, která se replikuje a přetrvává způsobem, jež odpovídá výchozím předpokladům (premisám anebo axiomům). Reformulace univerzální podoby darwinismu pak může být vyjádřena následovně (Dawkins 1983, [1989] 2003, Dennett 1995):
- Replikující se replikátory;
- Dědičnost vlastností replikátorů;
- Mutace replikátorů;
- Selekce replikátorů ve smyslu jejich úspěšnosti.
Univerzální darwinismus tak je zobecněním evolučních principů známé biologické skutečnosti do formy s mnohem širší platností, než představuje svět známých a neznámých živých forem.
Univerzální darwinismus ale není závazným pojmem pro vyjádření evoluce jako univerzálního principu anebo zákona. Někteří autoři ho odmítají pro jeho přílišný biologický nádech a nahrazují ho např. „obecně selekčními procesy“ (Hull 1988, 2001). Ve všech případech v podstatě jde o obecnou ideu evolučního procesu, kterou zastával Donald T. Campbell (1974).
Obecná idea evolučního procesu – Campbellovo pravidlo
Donald T. Campbell (1974) chápal evoluci v každé doméně, ať již biologické nebo společensko-kulturní, jako specifický a konkrétní projev působení obecných evolučních principů. Campbellův postup je tedy opačný. Nepostupuje zobecňováním a univerzalizací darwinovské biologické evoluce, ale vychází od univerzální platnosti evolučních principů, díky níž se stává každá konkrétní instance evolučních změn jejich nutným důsledkem. Campbell zdůrazňuje, že podobnosti mezi organickou a kulturní evolucí nejsou pouze náhodné, ale že jde o inherentní vlastnosti každého vyvíjejícího se systému, v kterém se vyskytují variace mezi replikovanými jednotkami a selekčně se zachovávají jen některé varianty na úkor ostatních. Proto také později chce Durham nazvat ideu obecného evolučního vývoje „Campbellovým pravidlem“, aby zdůraznil, že „analogie kulturních akumulací nejsou vztaženy k organické evoluci samé, ale spíše vzhledem k obecnému modelu evoluční změny […] pro který je organická evoluce pouze jednou z instancí“ (Durham 1992, 187). V tomto smyslu můžeme mluvit o „obecné evoluční perspektivě“ (Havlík 2008), jež nás nabádá k odhlédnutí od konkrétních případů evolučního procesu a vede nás k přehlédnutí všech evolučních procesů jako jednotlivých instancí obecného a univerzálního evolučního zákona.
Obecná forma selekčních procesů
Dalším příkladem, který se příliš nevzdaluje od biologické skutečnosti, je snaha vytvořit obecný evoluční koncept pro tři různé evoluční procesy - geneticky založenou selekci biologické evoluce, reakci imunitního systému na antigeny a operantní učení (Hull, Langman, Glenn 2001). Hull hovoří i o evolučním pojetí vědeckého poznání a naznačuje tak, že by pod takto zobecněné selekční procesy měla být zařazena i konceptuální změna ve vědě, která ale nemůže být založena geneticky (Hull 2001, 98). Všechny tyto konkrétní instance selekčních procesů však nelze jednoduše považovat za analogické a je třeba upozornit na jejich specifika a podstatné odlišnosti. Hull například říká, že „reakce imunitního systému na antigeny je selekční proces, který se odlišuje stejně tak radikálně od selekce založené na genech, jako se od ní odlišuje konceptuální změna ve vědě“ (Hull 2001, 98). Jde tedy o specifické a konkrétní případy selekce v různých oblastech skutečnosti a na základě různých jednotek (entit) selekce. Přestože jde o různé typy procesů, Hull nepochybuje, že všechny případy selekčních procesů jsou důsledkem jedné obecné formy selekčního procesu, o jehož analýzu mu jde. To, co spojuje všechny rozdílné selekční procesy do jednotné obecné koncepce, jsou tři základní prvky - replikace, variace a interakce (s okolním prostředím) -, jež musí být propojeny velmi specifickým způsobem. Například replikace musí být vystřídána interakcí s okolním prostředím, aby změny (variace), které nastaly při replikaci, měly během interakce rozdílný vliv. Na druhé straně ale existují i významné odlišnosti mezi těmito selekčními procesy, které se týkají především role organismu. Zde Hull, Langman a Glenn upozorňují na to, že pokud jde o interakce s okolním prostředím, hrají organismy v tradiční biologické evoluci centrální roli. Protože ale selekční procesy mohou nastávat na všech úrovních organizační hierarchie, je postavení organismu vůči nim rozdílné. Pokud např. jde o imunitní procesy, má organismus – na rozdíl od své centrální role v biologické evoluci – naopak funkci prostředí, ve kterém se interakce imunitního systému uskutečňují, a doba života organismu pak ohraničuje i existenci výsledků těchto selekčních procesů, protože změny dosažené v imunitním systému jedince se nepřenášejí na další organismy. Na druhé straně procesy operantního chování a učení nejsou ohraničeny tělem organismu, protože převážná většina interakcí, která vede ke změnám v nervovém systému organismu, je uskutečňována vůči okolí. Ani v tomto případě ale nedochází k přenosu výsledných změn selekčních procesů na potomky. K dalším důležitým aspektům patří odlišná role genů v jednotlivých selekčních procesech, shoda v iteračním charakteru všech procesů a konečně i neefektivnost a nákladnost těchto procesů, díky čemuž generují tyto procesy komplexitu a novost.
Instanciace evolučních procesů
Analogie v těchto případech tedy nejsou rozhodující. Mnohem vhodnější strategií je naopak analýza obecné formy selekčního procesu aplikovatelná stejně na všechny (možné) druhy selekce. Hull se v tomto ohledu odvolává na práce Stephena Toulmina (1972) a Donalda Campbella (1974). V této souvislosti pak požaduje, aby obecná analýza selekčního procesu byla na jedné straně „uspokojivě obecná“ (tj. aby nebyla jednak založena pouze na nějakém určitém typu selekce), ale na druhé straně nebyla tak obecná, aby se všechny přírodní procesy staly instancemi selekce. Instancemi musí být např. biologická evoluce, reakce imunitního systému nebo růst vzdělanosti, ale ne valící se koule nebo planety rotující kolem Slunce (Hull 2001, 98).
Úkolem tedy je nalézt univerzální formu souvislostí, jež by vyjadřovala podstatu selekčních procesů jako hlavního evolučního mechanismu a jež by byla schopná obsáhnout jako své jednotlivé případy jejich konkrétní instance. Viděli jsme, že Dawkins při svých úvahách o univerzálním darwinismu vychází ze dvou takových instancí - biologické evoluce a kulturní evoluce. Hull podporuje svůj koncept evoluce vědy (tj. instanci selekčních procesů ve vědeckém poznání) dalšími třemi již zmiňovanými instancemi - geneticky založenou selekcí biologické evoluce, reakcí imunitního systému na antigeny a operantním učením. Existují ale i další příklady selekčních procesů, které se objevují v oblasti genetických algoritmů, virtuálního života, počítačových virů i v pokusech uplatnit evoluční principy v kosmologii a kvantové teorii (Smolin 2004; Zurek 2003).
Nepochybně každá z takových instancí evolučního mechanismu selekčních procesů vykazuje určitá specifika a jedinečnost dané instanciace, jež mohou vést k zakrytí obecné souvislosti takových procesů. Úkolem obecné evoluční perspektivy tak je vyjádřit přesvědčení uniformity přírody v nejrůznějších oblastech skutečnosti, ať již jde o rozměrovou hierarchii (od mikro- k mega-rozměrům) nebo o úrovňovou hierarchii (mikro-makro) strukturní složitosti a komplexity. Nové a strukturně komplexní vzniká díky podobným principům a mechanismům. Obecná evoluční perspektiva tak předpokládá dostatečný odstup a abstrakci, jež umožňuje nahlédnutí těchto jednotlivých instancí jako důsledku obecných a uniformních vzorů (zákonů).
Principy univerzálního evolučního mechanismu
Někteří zastánci univerzálního darwinismu uvádí několik rozdílných způsobů, jak může být darwinovská evoluce konstruována (Plotkin 1993):
- Za prvé pomocí tří následujících principů:
- Existence fenotypových variací (individua se liší ve struktuře a funkcích);
- Variace vykazují odlišnou fitness/zdatnost (odlišné fenotypy mají různou schopnost přežití a reprodukce);
- Fitness/zdatnost je dědičná (znaky, které přispívají k fitness/zdatnosti rodičů budou přeneseny na potomky).
- Za druhé lze tento mechanismus vyjádřit vztahem replikátor – interaktor – rodokmen. To znamená, že existují replikátory, které interagují s okolím a vytvářejí přitom souvislou řadu generací (Lewontin 1970).
- Za třetí lze tento proces popsat jako hru slepých variací a selekčního uchování (Campbell 1974). Generují se variace bez předchozí znalosti jejich úspěšnosti a jsou selektivně uchovávány a stávají se původci další fáze variací.
- Konečně za čtvrté lze darwinovskou evoluci shrnout jako heuristiku „g–t–r“, kdy se systém generuje, testuje a regeneruje.
Obtíže jednotlivých modelů
Všechny tyto možnosti jistě vyjadřují důležité aspekty evolučního mechanismu, ale jen těžko lze předpokládat, že všechny stejně úspěšně vedou k rekonstrukci univerzálního evolučního mechanismu. Problémem jejich porovnání může být i odlišná terminologie, jež je důsledkem odlišných ontologických struktur.
- V prvním případě se tak např. mluví o fenotypových variacích, což je přímý odkaz k biologické formě interakcí, jež nemusí být na jiných úrovních skutečnosti vždy splněny.
- Ve druhém případě je užito distinkce replikátor – interaktor, jež je sice mnohem univerzálnější než distinkce genotyp – fenotyp, ale předpokládá další specifikace tohoto vztahu, především to, zda každý replikátor je interaktorem a každý interaktor replikátorem. Takové rozlišení vede k dalším důsledkům, pokud jde o úrovně selekce a základní jednotky evolučního mechanismu, jež jsou pro formulaci univerzálního evolučního mechanismu rozhodující.
- Hra slepých variací a selekčního uchování pak sice vystihuje rozhodující kroky v evolučním mechanismu, ale příliš dobře tento mechanismus nevyjasňuje. Slepost variací implicitně obsahuje neznalost úspěšnosti interakce s okolím a přímé vazby na přežití variací v těchto interakcích. Nejasný je také nositel těchto variací a způsob, jakým k variacím dochází. Obdobně také selekční uchování nejasně vyjadřuje, proč k uchování dochází a jakým způsobem to ovlivňuje další generaci variací. Souvislost variací a vytváření rodových linií je v tomto případě úplně zakryto.
- Poslední možnost je systémovým pohledem a vyjadřuje stavy systému v jednotlivých fázích evolučního mechanismu. Heuristika „g–t–r“ ale postrádá přesnější specifikaci, pokud jde o základní entity a jejich prostředí. Není jasné, zda regenerující se systém zahrnuje pouze měnící se entitu (replikátor), celou generaci entit (replikátorů), nebo dokonce i jejich okolí. Pro formulaci univerzálního evolučního mechanismu to opět jsou rozhodující aspekty.
Univerzální evoluci pravděpodobně není možné testovat přímo, ale jen prostřednictvím jejích konkrétních instanciací. Univerzální evoluční mechanismus tedy musí být průnikem všech těchto jednotlivých instanciací a musí splňovat požadavek dostatečných podmínek k realizaci evolučního mechanismu. Formulace univerzálního evolučního mechanismu potom musí mít podobu přírodního zákona, který se prosazuje vždy, když jsou splněny podmínky pro jeho uskutečnění. Konkrétní jednotlivosti a specifičnosti jednotlivých instanciací nejsou rozhodující, ale pouze dokreslují jednotlivé realizace evolučních procesů, a jsou tak důsledky rozmanitosti, s jakou se mohou evoluční procesy uskutečňovat v nejrůznějších úrovních skutečnosti a na základě různé komplexity entit a prostředí, jež se těchto procesů účastní.
Evoluční proces je nutným důsledkem přesně stanovitelných podmínek, který je nezávislý jak na substrátu v kterém vzniká, tak na hierarchické úrovni skutečnosti, v které se realizuje. Představuje nutné „vyrovnávání se“ entit se svým okolím, „začleňování se“ entit do svého okolí postupnými proměnami – adaptací. Tento obecný charakter evolučního procesu, kterým dochází i k evoluci biologických forem a jejich začleňování do daných přírodních prostorů a podmínek, se realizuje v nejrůznějších prostředích a na základě nejrůznějších entit, jež jsou jím „řízeny“ a organizovány.
Další možné interpretace axiomatizace evolučního mechanismu
Následující nebiologická interpretace axiomatizace evolučního mechanismu má demonstrovat, že takové interpretace jsou možné a že obdobným způsobem zobrazují relace a vztahy v reálném světě. V tomto případě jde o interpretaci takových počítačových instrukcí kódu, které vedou k sebereplikacím (tzv. počítačové viry).
Předpokladem takové interpretace jsou určitá zobecnění původně čistě biologické axiomatizace. Ta začíná vymezením vlastností souboru reprodukujících se organismů, na něž působí přírodní výběr. Každý soubor těchto vlastností je nazván biokosmem, primitivními termíny biokosmu jsou pak biologická entita a relace být rodičem (Williams 1970). Pro požadované zobecnění by postačilo nahradit biokosmos libovolným evolučním prostorem (navrhuji zde užít termínu evoprostor [evospace]) a biologickou entitu replikující se entitou nebo přímo replikátorem. Pak bychom mohli uvažovat také o evoprostoru počítačových virů, který by byl vymezen všemi prostředky umožňujícími replikace těchto kódů. I v tomto jednoduchém případu digitálního evoprostoru, který nám nyní postačuje jako příklad požadované interpretace, jsou splněny požadované axiomy relací být rodičem a být předchůdcem. Rodičovská relace je splněna při replikaci kódu a identifikace rodiče a potomka může být provedena v paměťovém prostoru digitálního počítače. Stejně tak předpokládáme, že jsou splněny i další axiomy darwinovské axiomatizace a že i triviálně digitální evoprostor může vést k evolučním mechanismům přírodního výběru, jak to naznačil Fred Cohen, který již v osmdesátých letech minulého století definoval pojem počítačového viru. Cohen upozorňuje, že většinou uvažujeme o evoluci počítačových virů prostřednictvím lidského programování, ale to neznamená, že se viry nemohou vyvíjet i jiným způsobem. Navrhuje mechanismus, jehož prostřednictvím by došlo k evoluci sebereplikujících se kódů. Uvažuje o programu, který používá pseudonáhodné hodnoty k pomalé změně vah jednotlivých funkcí programu z generace na generaci. Selekce a replikace nastává s pravděpodobností, která je vázána na efektivnost (úspěšnost) výsledné funkce programu. Pokud by byly parametry variovány vzhledem k úspěšnosti programu vhodným způsobem, „druhy“ dostupných virů se budou „vyvíjet“ k mnohem efektivnějšímu souboru programů (srov. Cohen 1991).
Genetické algoritmy, evoluční a genetické programování
Vezměme jako příklad oblast genetických algoritmů GA (např. Holland 1975), evolučního programování ES (např. Rechenberg 1971) a genetického programování GP (např. Koza 1992). Všechny tyto přístupy k programování vycházejí z inspirace evolučními procesy v biologii. Uvádějí několik obecných principů nebo podmínek darwinovské evoluce a pak abstrahují od biologických entit, genů, organismů, zdatnosti, mutace, přežití atd. k jejich počítačovým reprezentacím. Například John R. Koza uvádí čtyři nutné podmínky k tomu, aby v přírodě nastal evoluční proces (Koza 1992, 17):
- Entita, jež má schopnost se reprodukovat.
- Existence populace takových sebe-reprodukujících se entit.
- Existence rozdílů (variet) mezi sebe-replikujícími se entitami.
- Určité odlišnosti ve schopnosti přežít v prostředí jsou spojeny s varietami.
Vidíme, že uvedené podmínky jsou až na drobné formulační odchylky blízké výše uvedené možnosti reformulace univerzálního darwinismu, provedené na základě rekonstrukce darwinovských podmínek reprodukce, dědičnosti, variability a selekce. Vzhledem k biologické evoluci by sice mohly být vzneseny námitky vzhledem k tomu, že není explicitně zdůrazněn požadavek nadprodukce potomků a omezenosti zdrojů, aby tak mohl začít fungovat přírodní výběr, nebo že není explicitně zdůrazněn požadavek dědičnosti. Problematizováno by tak mohlo být implicitní obsažení těchto předpokladů v první a poslední podmínce. To však není pro náš příklad rozhodující. Důležité je, jak pokračujeme, máme-li k dispozici (více či méně) vhodnou reformulaci podmínek biologické evoluce. Formulujeme základní cyklus evolučních algoritmů, podle kterého fungují v zásadě všechny GA (např. podle T. Weise 2009, 96) (obr. 1):
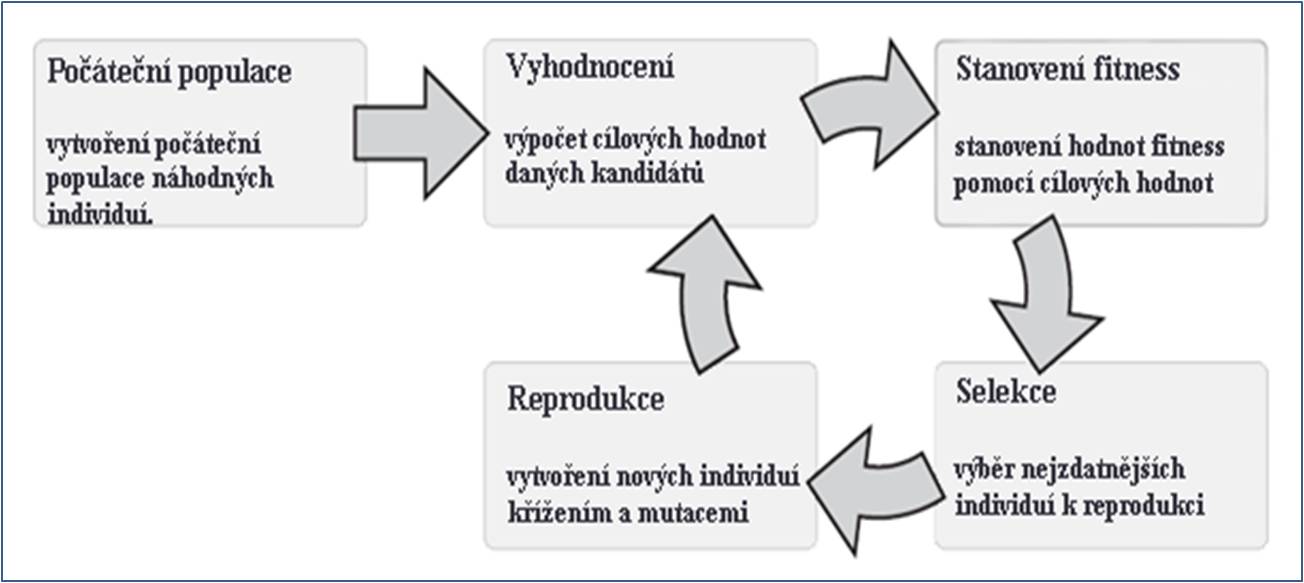
Obr. 1: Základní cyklus genetických algoritmů
Obrázek 1 ukazuje základní cyklus pro všechny varianty GA, a může tak být chápán jako zobecnění nebo obecný princip všech jednotlivých instancí GA. (Pozn.: Jeho modifikací bychom mohli dostat další zobecnění, jež by zahrnovalo i třídu genetického programování, GP, jež se liší od třídy GA tím, že se vztahuje i na samotné tělo programového kódu, který je v evolučním cyklu „náhodně“ modifikován tak dlouho dokud jeho funkce nevede k požadovaným vlastnostem.) Cyklus genetických algoritmů se od biologického evolučního procesu liší v tom, že obsahuje vstupní a výstupní kroky, které slouží jednak pro iniciaci a jednak pro přerušení procesu. Smyslem GA není simulace evolučního procesu, ale jeho využití pro hledání nějakých výpočetně časově náročných řešení (např. minima či maxima nějaké funkce). V tomto smyslu lze říci, že procesy GA jsou cílesměrné, protože jsou využity pro hledání co nejlepšího řešení mezi ostatními. To by zdánlivě mohlo vést k názoru, že nemají nic společného s biologickou evolucí, která cílesměrná není. Teleologie je v případě biologické evoluce přírodním výběrem jednoznačně popřena a vysvětlení zdánlivě záměrných znaků a vlastností organismů je plně dosaženo právě mimo předpoklad záměrného a cílevědomého působení. Takový argument je však mylný. Uspořádání GA nijak nenarušuje obecný evoluční princip, který tvoří jádro GA, a plně využívá možností evolučního procesu jako takového. Pouze doplňuje počáteční hodnoty (podmínky) jeho fungování a testuje v jistém smyslu fitness jedinců každé populace a po dosažení předem stanovené cílové podmínky pak evoluční cyklus ukončuje.
Názorněji je to vidět na příkladu algoritmizace takové obecné struktury pro hledání určitého optima, jež může vypadat následovně (viz Obitko 2010):
2. [Start] Generuje se náhodná populace n chromozomů.
3. [Fitness] Vyhodnotí se fitness f(x) každého chromozomu x v populaci.
4. [Nová populace] Vytvoření nové populace opakováním následujících kroků.
1. [Selekce] Vyberou se dva rodičovské chromozomy z populace podle jejich fitness (čím lepší fitness, tím větší pravděpodobnost že bude vybrána).
2. [Křížení] Na základě pravděpodobnosti křížení se vytváří noví potomci. Nedojde-li ke křížení, potomci jsou přesné kopie rodičů.
3. [Mutace] Na základě pravděpodobnosti mutace se změní noví potomci v každé pozici chromozomu.
4. [Přijetí] Noví potomci se umístí do nové populace.
5. [Nahrazení] Nově vygenerovaná populace se užije pro další běh algoritmu.
6. [Test] Je-li dosaženo cílové podmínky, stop, a vrať nejlepší řešení ve stávající populaci.
7. [Loop] Pokračuj bodem 2.
I v tomto případě můžeme rozlišit tři již zmiňované základní prvky - replikace, variace a interakce -, které jsou pro úspěšný běh takového optimalizujícího algoritmu podstatné. Přestože v tomto případě jde o složitější model, jež využívá křížení a mutací ke vzniku nové populace, je jasné, jakým způsobem je v tomto případě replikace a variace implementována. Méně evidentní je ale třetí prvek, tj. interakce. I ten je ale v algoritmu obsažen, a to ve vyhodnocení fitness chromozomu a pravděpodobnosti, s jakou bude daný chromozom selektován do další populace.
Evoluce jako algoritmický proces
Příklad GA poukazuje k další důležité skutečnosti, která má dva různé důsledky. Zaprvé ukazuje, že evoluční proces je nutné chápat jako algoritmický děj. V případě organické evoluce sice nevystupuje tento rys nějak výrazněji do popředí, ale na druhé straně také organická evoluce neobsahuje nic, co by zmiňované algoritmizaci bránilo. Tento rys evoluce byl zdůrazněn Calvinem až v 80. letech minulého století (Calvin 1987), když použil termín Darwinův stroj, aby tím vyjádřil podstatu evoluce jako algoritmického procesu. Analogicky jsou používány podobné termíny - např. Turingův stroj nebo von Neumannův stroj – ke zdůraznění algoritmičnosti operací, jež se v daných mechanismech provádějí. Později také Plotkin (1993) přijímá stanovisko univerzálního darwinismu a definuje tyto univerzální evoluční procesy jako Darwinův stroj:
- „[…] ačkoli lze dospět k vysvětlení transformací živých systémů evolucí, která působí mezi organismy, procesy, jež vedou k evoluci, jsou vždy totožné. Vycházejí z univerzálních procesů variace, odlišné zdatnosti (fitness) a dědičnosti, přenosu vybraných variant a jejich kombinací s novými variantami, jež vedou k univerzálnímu darwinismu a definují Darwinův stroj.“ (Plotkin 1993, 86; kurzíva v originále).
Jednoduchost a komplexita
Druhým důsledkem, který těsně souvisí s pojetím evoluce jako algoritmického procesu, je vazba mezi rozmanitostí a komplexitou organického života a jednoduchostí a opakovatelností základních kroků zmiňovaného evolučního algoritmu. V univerzálním případě pak odhlédneme od organické instance evoluce a přisoudíme tuto schopnost univerzálním evolučním principům. Algoritmické mechanismy, které se dějí na určité primární úrovni jednoduchým a opakovatelným způsobem, vedou na sekundární úrovni k rozmanitosti, komplexitě a organizované uspořádanosti a složitosti. Tímto způsobem je pak možné popsat Darwinův stroj jako fungování algoritmického procesu, který je ve svém zásadním principu jednoduchý a opakovatelný, ale který právě díky tomu také vede ke vzniku organizované uspořádanosti (např. ke vzniku takových orgánů, jako je oko).
Případ GA ale ukazuje, že v daném izolovaném evoprostoru, pokud bychom nechali proběhnout evoluční cyklus do té doby, než by se hledaný organismus v populaci objevil. (Pozn.: Podobné experimenty se provádějí virtuálně např. užitím GA k evoluci virtuálních tvorů schopných realistického pohybu ve fyzikálně simulovaném 3D prostředí. Viz např. http://www.spiderland.org/screensaver.) Darwinův stroj funguje v obou těchto procesech identicky. Principiálně jde o různé realizace univerzálního evolučního vzoru. Vzhledem ke zmiňovaným sekundárním důsledkům realizace tohoto vzoru, jež spočívá v rozmanitosti a variabilitě, bychom mohli hovořit o tvořivosti přírodních procesů, aniž bychom byli nuceni odvolávat se na teleonomii, plánovitost a záměrnost.
Předpoklady reformulace
Jedním z problémů univerzální reformulace evoluce v rámci univerzálního darwinismu je, že R. Dawkins, který navrhuje toto zobecnění, trvá na tom, že pokud jde o biologickou evoluci, je rozhodující distinkce mezi replikátorem a vehiklem. Tím poněkud komplikuje obecné pojetí replikátoru.
Replikátory germ-line a dead-end
Podle Dawkinse lze klasifikovat replikátory dvěma nezávislými způsoby. Jsou to buď replikátory germ-line, nebo replikátory dead-end a jako takové jsou buď aktivní, nebo pasivní. Germ-line replikátor je potenciální předchůdce nekonečně dlouhé linie svých potomků, zatímco dead-end replikátor, který se sice také může replikovat, vytváří jen konečnou řadu svých potomků a není potenciálním předchůdcem jejich nekonečné linie. Aktivní replikátor je každý replikátor, jehož povaha má vliv na pravděpodobnost, že bude replikován. Pasivní replikátor nemá vliv na pravděpodobnost své replikace. Ze všech čtyř možností, které kombinačně přicházejí do úvahy, je nejdůležitější jednotkou aktivní germ-line replikátor, protože ať už se nalézá kdekoli ve vesmíru, může se stát základem přírodního výběru, a tedy evoluce (Dawkins 1982, 83–84). Gen, který má fenotypové vyjádření, tedy je aktivním replikátorem. Na rozdíl od toho některé části DNA, které nikdy nejsou transkribovány a nemají žádné fenotypové vyjádření, jsou pasivními replikátory. Za pasivní replikátor tak může být považována část DNA pouze tehdy, jestliže její povaha nemá absolutně žádný vliv na pravděpodobnost její replikace (Dawkins 1982, 83; 1982a, 162). Problémem ovšem je, že předpoklad o částech DNA, jež nejsou nikdy transkribovány a nemají žádný vliv na pravděpodobnost své replikace, je zatím pouhou domněnkou, protože ještě nejsou přesně známy všechny funkce částí DNA a jejich role v prostorovém uspořádání DNA jako komplexního celku. Navíc se v rámci výzkumů tzv. Evo-Devo stále více ukazuje, že právě ty části DNA, které nenesou přímou fenotypovou informaci, obsahují řídící kódy pro vývojový proces individuálního organismu, a jsou možným předmětem mutací a evolučních změn.
Dawkinsův genocentrismus je na základě distinkce replikátor – vehikl poněkud matoucí z hlediska snahy o vyjádření univerzálního evolučního mechanismu. Je totiž nutné, aby obecné principy postihly mnohem širší třídu jevů, než je biologická evoluce a než je její poměrně specifické genocentrické Dawkinsovo pojetí. Dawkins se sice snaží zobecnit a univerzalizovat jednotku selekce, když transponuje gen na replikátor jako na libovolnou entitu, z níž se vytvářejí kopie, ale v zásadě jeho pojetí nepočítá s jinými případy replikátoru, než jsou
- gen jako jednotka organické evoluce,
- hypoteticky jakýkoli jiný negenetický základ života v kosmu,
- mem jako jednotka kulturní evoluce.
Z toho důvodu je jeho pojetí replikátoru oprávněné, neboť všechny tři případy splňují požadavek fenotypového vyjádření. Geny budují své vehikly (fenotypy), jež jim zajišťují jejich přežití, memy se mohou seskupovat do memplexů a také se „fenotypizovat“ prostředky, jež jim zajišťují jejich další přežití. Na druhé straně ale existují procesy, které obsahují entity – replikátory, které vykazují evoluční chování přes to, že nemají fenotypové vyjádření.
Negenocetrické evoluční procesy
Příkladem takových typů procesů jsou například reakce imunitního systému na antigeny, kde se mutované geny selektují okamžitým testováním jejich proteinového produktu. V tomto případě se vztah genu a jeho proteinového produktu pravděpodobně nedá považovat za identický se vztahem mezi genotypem a fenotypem, přestože selekce genů imunitního systému probíhá na úrovni jejich proteinových produktů. Obdobně také selekce virů (bakteriofágů) probíhá selekčními mechanismy, ale díky specifickému horizontálnímu genovému transferu (HGT) vykazují bakteriofágy mozaicismus a jejich současná rozdílnost je pravděpodobně způsobena kombinací vertikální a horizontální evoluce. „Bakteriofágy jsou vyvíjející se agenti (činitelé), ale vyvíjejí se, aniž by vytvářely druhy; vytvářejí neuvěřitelnou rozmanitost biologických agentů, kteří mají přístup k enormně rozsáhlému společnému genofondu.“ (Hendrix et al. 1999).
Díky takovému biologickému uspořádání je třeba znovu uvážit, jak pracuje přírodní výběr. Bakteriofágy jsou důkazem, že přírodní výběr nemusí nutně působit na populace a nemusí vést nutně ke vzniku odlišných druhů. Bakteriofágy jsou díky svému mozaicismu individui složenými ze spletitých chimérických genomů a fenotypů, které neodpovídají žádné určité evoluční historii. Zdá se tedy, že se nejhojnější biologičtí aktéři vyvíjejí, aniž by formovali biologické druhy (Morgan and Pitts 2008). Ani v tomto případě není tedy plně dodržena distinkce mezi replikátorem a vehiklem v Dawkinsově smyslu. Nahrazení biologického organismu obecnou entitou (agentem, replikátorem) v principu variability, může být tedy provedeno pouze tak, že takto zobecněná entita, jež se účastní evolučního procesu, nebude zatížena distinkcí replikátor – vehikl. Neznamená to, že je třeba odmítnout tuto distinkci samotnou, ale pouze přijmout tu skutečnost, že pro formulaci univerzálního evolučního mechanismu není podstatná. Na úrovni biologických druhů je sice významnou charakteristikou průběhu evolučního procesu, což ale neznamená, že musí být takto explicitně přítomna i na jiných úrovních, kde dochází k instancím univerzální evoluce. Distinkce replikátor – vehikl (genotyp – fenotyp) může být mnohem slabší (imunitní systém) nebo mnohem komplikovanější (bakteriofágy), případně může zcela chybět (genetické algoritmy).
Základní jednotka selekčních mechanismů přírodního výběru
Ernst Mayr předpokládá, že nejnižší úroveň organizace živé hmoty, která prochází evolučními změnami, je populace. Jedinci v genetickém smyslu neprocházejí evolučními změnami, a proto můžeme evoluci nejvýstižněji charakterizovat jako genetickou změnu příslušníků každé populace z generace na generaci (Mayr 2009, 113). Co je však základní jednotkou selekčních mechanismů přírodního výběru? V této souvislosti lze rozlišit několik základních strategií, které byly a jsou v diskusích zastávány:
- Replikátory. Dawkins předpokládá, že jednotkou selekce je pouze replikátor. Popírá ale, že by mohla existovat hierarchie replikátorů odpovídající hierarchii úrovní organizace života (prebiotická molekula, gen, organismus, populace, druh, ekosystém) a vyžaduje, aby v biologické oblasti byla role replikátoru přisouzena pouze genu. (Pozn.: S tou výhradou, že přesné určení replikátoru jako jednotky není důležité, neboť může jít o libovolně dlouhé úseky DNA, a díky tomu lze tedy připustit i jistou hierarchii replikátorů. Taková hierarchie je však omezena pouze na gen, díky jeho specifickému postavení v evolučním procesu.)
- Vehikly. Wilson a Sober přijímají Dawkinsovu distinkci mezi replikátorem a vehiklem, ale na rozdíl od něho naopak považují vehikly za jediné jednotky selekce, protože podle jejich názoru selekční tlak působí pouze na fenotypy.
- Žádné jednotky selekce neexistují. David Hull preferuje interpretaci, podle které zde nejsou žádné jednotky selekce, protože selekce sestává ze dvou podprocesů, z replikace a z interakce, a selekce sama je pak mezihrou v těchto dvou podprocesech. Proto je podle Hulla zavádějící mluvit o jednotce selekčního procesu.
- Hierarchie replikátorů. V tomto případě se předpokládá, že replikátor je univerzálním označením, jež je aplikovatelné na různých úrovních organizace živého. Jako replikátor potom může být chápán nejen gen, ale i organismus, populace, druh, vyšší taxonomické jednotky nebo celý ekosystém. Například Brandon takto navrhuje duální hierarchii interaktorů a replikátorů odvozenou z jednotlivých úrovní biologické domény.
- Vývojový proces nebo životní cyklus. Nejkomplexnější pojetí jednotky selekčního procesu v rámci myšlenkového proudu tzv. systémově-vývojové teorie. Zdůrazňuje se zde komplexita probíhajících procesů (např. interakce genu s okolím při replikaci), která má být důvodem pro neodlišitelnost jednodušších entit v selekčním procesu, než je jeden vývojový nebo životní cyklus (Oyama, Griffiths, Gray 2001).
Základní jednotka selekce
Podobné je to i v případě sporu, zda je jednotkou selekce replikátor, nebo vehikl, případně genotyp, nebo fenotyp. Je třeba si uvědomit, zda je v případě otázky jednotek selekce tato základní distinkce rozhodující. Posuzujeme-li tuto otázku výhradně z pohledu biologické evoluce, pak nás výrazný rozdíl mezi genotypem a fenotypem může zmást. V kontextu univerzální evoluce ale nemusí být tento rozdíl tak výrazný, nebo dokonce nemusí být vůbec rozlišitelný, a díky tomu není pro pojetí univerzálního evolučního mechanismu rozhodující. Diskuse, zda jednotkou selekčních změn je gen (replikátor), nebo fenotyp (vehikl), nemůže nijak podstatně změnit tu skutečnost, že mezi genem a fenotypem existuje specifická vazba proměny informací uložených v genu prostřednictvím vývoje organismu a jeho rozsáhlých fenotypových projevů v daných podmínkách, jež rozhodují o uchování a další replikaci těch informací, které se v daných podmínkách ukázaly jako úspěšnější. Vyčleňovat z těchto souvislostí jedinou komponentu jako jednotku selekce se nezdá být konstruktivní. Fenotyp jako základní jednotka selekce je nemyslitelná bez její ontogenetické vazby na genotyp a naopak genotyp jako základní jednotka selekce zase nemůže být přímo selektována přírodním výběrem. S touto provázaností souhlasí i Dawkins a přes svůj silný požadavek genocentrismu se dokonce i omlouvá za to, že jeho fráze „selekce replikátorů“ byla nesprávně pochopena tak, že by geny nebo jiné replikátory skutečně čelily přírodnímu výběru, a že je naopak třeba uvážit selekci vehiklů, protože replikátory nejsou přírodnímu výběru přímo viditelné (Dawkins 1982a, 2–6).
Někteří autoři předpokládají, že pojem replikátoru nemůže být vyhrazen pouze genům a že Dawkinsův genocentrismus je příliš restriktivní. Na druhé straně je však otázka, zda lze připustit Brandonovu (Brandon 1998) duální hierarchii interaktorů a replikátorů odvozenou z jednotlivých úrovní organizace života v rámci biologické domény. Existuje hierarchie replikátorů v biologické říši, nebo zde jsou fundamentální důvody pro přisouzení role replikátoru pouze genu?
Hull je přesvědčen, že diskuse o základní jednotce selekce jsou pochybné, protože žádné takové jednotky neexistují. Hull chápe selekci jako mezihru mezi dvěma fundamentálními procesy, mezi replikací a interakcí. Selekce sama jako mezihra nemá žádné jednotky, které by byly selektovány, ale procesy replikace i interakce je mají - replikátor a interaktor. Díky tomu je klasická organizační hierarchie gen - organismus - druh nepřirozená a je třeba ji nahradit jinou organizační hierarchií. „Náležitými úrovněmi nejsou geny, organismy a druhy, jak se tradičně předpokládá, ale replikátory, interaktory a rody (lineages).“ (Hull 2001, 21).
Hull vychází z Dawkinsovy snahy nahradit pojem genu obecnějším termínem replikátoru, ale zároveň ukazuje, v čem je Dawkinsovo pojetí nedostatečné. Dawkins definuje replikátor jako entitu, jejíž struktura zůstává nedotčena v následných replikacích. Podle Hulla je ale pouhé zachování identity struktury replikátoru pro selekční proces nedostatečné. Za rozhodující považuje identitu v rodové linii (tzv. IBD – identical by descent, tj. „pocházející ze společného předka“). (Pozn.: Obecně se rozlišují alely identické v rodové linii – IBD, identical by descent, tj. „pocházející od společného předka“, a alely identické de facto – IBS, identical by state). Alela IBD je zároveň alelou IBS, opačně to ale neplatí. Vzhledem k důležitosti alel IBD je tato vlastnost nově definována vzhledem k faktu dědičnosti tak, že dvě nebo více alel jsou IBD, jestliže byly zděděny ze stejné alely bez rekombinace.) Kromě toho se Hull pozastavuje i nad samotným požadavkem identity, protože se zdá být příliš omezující pro individuaci replikátoru, vzhledem k mnoha mutacím, které při replikaci nastávají.
IBS a IBD
Faktem je, že termínu identita se užívá nepřesně a je vhodné vyjasnit, v jakém smyslu o identitu jde, a v jakém ne. Poněkud matoucím dojmem mohou působit zmiňované dva typy identičnosti alel IBS (identical by state) a IBD (identical by descent) a závěr, že alela IBD je zároveň alelou IBS, přičemž opačně to neplatí. IBS zaručuje pouze statickou identitu alely, která tak nemusí pocházet ze stejné rodové linie, což je splněno pouze v případě identity IBD. Vtip však spočívá v tom, jak se identita (ať již IBS nebo IBD) identifikuje. Obecným způsobem, jak takové určení provést, je použít jednonukleotidový polymorfismus (SNP – single nucleotide polymorphism), při kterém jsou porovnávány odchylky individuálních nukleotidů v sekvenci DNA. Pokud je pozorována shodnost dostatečného počtu SNP ve dvou alelách, pak jsou takové alely považovány za IBD, ačkoli to nezaručuje, že jsou skutečně identické! A dokonce i když jsou skutečně odvozeny z jedné alely jako jejich předka, mohou vykazovat odlišné mutace, které vznikají během replikace, a takové alely tedy nejsou identické ve striktním smyslu slova. Proto Hull zdůrazňuje nutnost IBD včetně variací, ke kterým během replikace nutně dochází. Domnívá se také, že připustit variace, které obecně mají minimální vliv na funkci genu, takže v rámci replikace je možné považovat ho stále za „ten samý“ gen, podstatně nemění původní smysl Williamsova a Dawkinsova pojetí, ale opuštění požadavku identičnosti v rámci rodové linie ano (Hull 2001, 22).
Replikace a interakce
Hull zdůrazňuje, že pro adekvátní pochopení selekce není pojem replikátoru postačující podmínkou. Upozorňuje na interakci jako další nutnou a podstatnou podmínku evolučního procesu. V Hullově pojetí evolučního mechanismu tak jsou odlišitelné dva základní procesy - proces replikace a proces interakce. Oba tyto procesy také mají své základní entity. Je-li replikátor základní jednotkou procesu replikace, pak je interaktor základní jednotkou procesu interakce. Hull definuje interaktor jako entitu, která „interaguje jako soudržný celek se svým okolím takovým způsobem, že tato interakce zapříčiňuje odlišnost replikací“ (Hull 2001, 22). Díky těmto dvěma procesům, replikaci a interakci, pak lze univerzálně definovat selekci jako „každý proces, v kterém různorodá vyhynutí (extinction) a rozšíření (proliferation) interaktorů zapříčiňují odlišná uchovávání replikátorů, které je produkují“ (Hull 2001, 22).
Hull zdůrazňuje, že pojem interaktoru je definován s odpovídající obecností a není omezen na jednu úroveň hierarchie, obdobně jako replikátor. Distinkce replikátor – interaktor je tedy distinkcí vztahující se k rolím, jaké daná entita v daném procesu naplňuje, a tatáž entita může hrát obě role. Například gen je nejen replikátorem, ale i interaktorem. Interaguje se svým okolím a může tím ovlivnit četnost replikací, podobně jako interagují se svým okolím organismy, ale i vyšší, inkluzivnější entity biologické hierarchie (kolonie, roje a podobné skupiny). Hull se tak snaží nahradit tradiční organizační hierarchii hierarchií, jejíž úrovně jsou vymezeny v termínech evolučního procesu samého (Hull 2001, 23).
Splňuje-li nějaká entita požadavek, aby interagovala jako soudržný celek se svým okolím tak, že má vliv na replikace, pak je interaktorem na libovolné úrovni biologické hierarchie. Na druhé straně otázka stejné univerzality replikátoru je problematičtější. Hull nejprve zdůrazňuje, že podmínkou toho, aby nějaká entita fungovala jako replikátor, je, že taková entita musí mít strukturu a musí být schopna předávat tuto strukturu v následných replikacích. Především je nutné, aby replikátor předával svou schopnost replikovat se na své repliky. „Ačkoli geny jsou dobře adaptovány pro funkci replikátoru, nevyplývá z takové definice, že geny jsou jediné replikátory.“ (Hull 2001, 110).
Otázka, zda by i jiné interaktory mohly sehrávat roli replikátorů, je podstatná. Přestože Hull původně s touto možností počítal, ale později ji díky diskusím o jednotkách selekce zmírnil, neznamená to, že by otázku replikátorů vyřešil v těch instancích evolučního procesu, o které se sám opírá, tj. např. v reakcích imunitního systému, operantního učení a konceptuální evoluce. Jde-li skutečně o instance selekčních procesů, pak by replikátory a interaktory měly být rozlišitelné i v těchto případech. Jinak by totiž distinkce replikátor – interaktor byla specifická pouze pro biologický případ selekčních mechanismů a nemohla by být znakem obecných evolučně-selekčních mechanismů.
Rodové linie
Pro výsledky replikace a interakce zavádí Hull obecný termín rod (lineage). Je to entita rozložená v čase, uspořádaná rodová linie následných generací – rod či rodokmen. Definuje ho jako „entit[u], která přetrvává neurčitou dobu v čase, buď v tom samém, nebo změněném stavu, jako výsledek replikace“ (Hull 2001, 110). Rod je tak historická entita formovaná sekvencemi replikací, prostoročasový řetězec předchůdců a následníků. Pro vznik rodové linie je rozhodující proces replikace. Proces interakce sice není pro rodovou linii podstatný, ale ke změnám rodu v čase dochází díky mezihře mezi replikací a interakcí. Hull předpokládá, že jak geny, tak organismy vytvářejí rody, zatímco druhy ne. Druhy neformují rodové linie, ale samy jsou rodovými liniemi (Hull 2001, 111).
Organismy jako replikátory
V uvedeném smyslu mluvíme o organismech jako o fundamentálních rozlišitelných entitách organického světa, které se reprodukují a vytvářejí struktury rodových linií předchůdců a následníků charakterizovaných specifickým vztahem být při reprodukci rodičem svých potomků. Rodové linie jsou entitami rozloženými v čase. Jsou propojeny uvedenou základní vazbou a rozkládají se od minulosti do budoucnosti. V tomto čistě jevovém, fenomenologickém smyslu tedy považujeme i organismy za replikátory. Dawkinsova výhrada vůči tomuto návrhu – tj. že replikátor musí být schopen přenášet svou strukturu do dalších replikací, a na úrovni organismu k replikaci v tomto smyslu nedochází, protože získané vlastnosti se nedědí (jinými slovy, evoluce není lamarckovská) –, zde neobstojí z toho důvodu, že nepožadujeme, aby organismus ve svých potomcích replikoval všechny své vlastnosti (tedy i ty získané). Plně totiž postačí, že jsou přeneseny některé z vlastností organismu, tj. ty genetické – a v případě sexuálně se reprodukujících organismů ještě bereme v potaz specifičnost sexuální reprodukce.
Hierarchie replikátorů?
Obdobně bychom mohli hledat i další případy replikátorů na vyšších hierarchických úrovních skutečnosti. Vše, co je schopno předat svou strukturu prostřednictvím svých replikací, je možné na dané fenomenologické (jevové) úrovni chápat jako replikátor. Samozřejmě tím nijak nepopíráme možnost, že existují i jiné možnosti vysvětlení a predikcí, které nepracují pouze na dané fenomenologické úrovni, ale zaměřují se na mikroskopické mechanismy (tj. mechanismy nižších úrovní), jež stojí v základech jevových forem dané úrovně. Z fenomenologické perspektivy dokonce můžeme označit za replikátor i druh. Nehledě na problematičnost jeho biologického vymezení, druh může být chápán jako určitá struktura organismů, jež jsou vázány vztahy následnosti a reprodukce v čase. Vzhledem ke stromové struktuře organických bytostí je časové vymezení druhu vágní, nicméně struktura mezi žijícími a již nežijícími příslušníky druhu je dána, stejně jako je predikovatelná struktura mezi žijícími příslušníky druhu a jejich potomky. Proto můžeme druh chápat jako relativně samostatnou entitu, jež je schopna reprodukovat danou strukturu v dalších generacích.
Hypotetické replikátory?
Pravděpodobně by bylo možné hovořit o replikátorech i na vyšších hierarchických úrovních. Pokud by došlo k replikaci organického světa planety Země na jiných vhodných planetách jiných slunečních soustav, mohli bychom dokonce celý tento organický svět jako celek pokládat za replikátor – pokud bychom předpokládali, za prvé, že by byla replikována daná struktura, a za druhé, že organický svět Země by měl přímý vliv na svou replikaci.
Reformulované principy obecného evolučního procesu
Souhrnně lze tedy navrhnout následující reformulaci základních principů obecně evolučních procesů (Havlík 2012):
- Časová posloupnost rodově provázaných generací variabilních entit, jež se vzájemně liší některými znaky.
- V každé generaci se replikuje takové množství entit, které nemůže být v daném prostředí uchováno. O jejich uchování a další reprodukci rozhoduje jejich interakce s prostředím.
- V rámci dané generace entit existují takové rozdíly mezi jejich vlastnostmi, jež mají přímý vliv na možnost uchování těchto entit a jejich další replikace.
- Většina vlastností entit v dané generaci se dědí na jejich potomky při replikaci.
Uvedené principy jsou dostatečně univerzální, aby pokryly všechny instance evolučních procesů, nehledě na entity, jež v nich vystupují, ani nehledě na hierarchickou úroveň, v níž se evoluční mechanismus realizuje.
Uvedená formulace je sice úplná, ale je jen jednou z možných formulací podobných principů, jež vyjadřují všechny nutné aspekty evolučního procesu – existenci variabilních entit, existenci selekčního tlaku, přímý vliv vlastností entit na jejich uchování a dědičnost.
Konečně také takto obecně formulované principy evoluce ukazují, že evoluční proces je nutné chápat jako obecný a nutný zákon, který se realizuje vždy, když jsou splněny všechny podmínky. Jinými slovy, je-li iniciován proces evoluce, nemůže pokračovat jiným způsobem než tak, jak jej popisuje tato univerzální formulace. Na druhé straně můžeme díky zákonitému charakteru evolučního procesu, stejně tak jako v jiných obdobných případech, pomocí takového zákona nejen vysvětlovat výsledky evoluce, ale můžeme také predikovat jeho budoucí výsledky.
Možnosti predikce výsledků evoluce
Upozornili jsme již na to, že predikce výsledků neznamená v tomto případě přesné určení např. druhových forem, které se v té či oné době objeví, protože novost znaků, tvarů a vlastností je závislá jak na niterných genetických mechanismech (jež jsou předmětem výzkumů nové vědy, tzv. Evo-Devo), tak na mnoha různých vlivech prostředí, ve kterém se evoluční mechanismus realizuje.
Predikovat však můžeme mnoho dalších podrobností výsledků evolučního procesu, které se týkají nejen jeho obecných souvislostí, ale i docela konkrétních předpovědí. Víme například, že organická evoluce vede k rozmanitosti a diverzifikovanosti a že postupná adaptace vede k tomu, že všechna spektra dostupného evolučního prostoru jsou zaplněna vhodnými formami. Zároveň víme, že existují určité fyzikální podmínky, v nichž se evoluce realizuje, a ty stanovují možný prostor evolučních forem. Jsou tak stanoveny možné způsoby pohybu a získávání energie. Nevíme sice přesně, jaké formy se v evolučním prostoru objeví, ale můžeme odhadnout dostatek jejich základních vlastností a kontur, ať už jde o způsoby pohybu v daném fyzikálním prostředí (chůze, plazení, let, plavání), další rozdělení organismů podle způsobů přijímání potravy (producenti, konzumenti, dekompozitoři), podle typu potravy (masožravci, býložravci, všežravci) anebo potravních řetězců (fytofág, zoofág, saprofág, koprofág, nekrofág) atd.
Lze předpokládat, že obdobným způsobem lze postupovat i v jiných instancích evolučních procesů. Nemůžeme sice přesně určit, jaké evoluční formy se objeví, ale můžeme predikovat dostatek podrobností a základních charakteristik pro daný evoprostor. Např. mluví-li se – většinou pouze v přeneseném významu – o evoluci počítačových virů, pak je možné ukázat, že skutečně jde o instanci obecných evolučních mechanismů, a pro tento evoprostor lze také predikovat jeho základní vlastnosti a kontury. Tato skutečnost je evidentní především při přenosu sebereplikujících se počítačových kódů, kdy jsou postupně využívány všechny dostupné cesty a způsoby, které infikaci umožňují. Způsoby přenosu a infikace jsou přitom závislé na rozvoji struktur počítačových technologií, které se postupně objevují, a také na všech evoprostorech vyšších vrstev virtuálního světa, jež se postupně vrství na základních, elementárních strukturách.
Obdobné predikce můžeme provést i v ostatních případech realizace obecných evolučních procesů, pro které jsme se pokusili nalézt jejich univerzální principy. Ty jsou podle našeho názoru základním předpokladem evoluční perspektivy, z níž je třeba pohlížet na většinu oblastí vyvíjejícího se a hierarchizovaného světa včetně vědomí a vědění, jež umožňuje evoluční sebereflexi.
Použitá literatura a literatura k dalšímu studiu
Barros, D. B. (2008). Natural Selection as a Mechanism. Philosophy of Science, vol. 75, 2008, s. 306-322.
Brandon, R. N. (1978). Evolution. Philosophy of Science 45, s. 96–109.
Brandon, R. N. (1998). The Levels of Selection: A Hierarchy of Interactors. In: D. L. Hull – M. Ruse (eds.). The Philosophy of Biology. Oxford, Oxford University Press, s. 176–197.
Calvin, W. H. (1987). The Brain as a Darwin Machine. Nature 330, 5/9/1987. Dostupné na: http://williamcalvin.com/1980s/1987Nature.pdf.
Campbell, D. T. (1974). Evolutionary Epistemology. In: P. A. Schilpp (ed.). The philosophy of Karl R. Popper, LaSalle, Ill., Open Court, s. 412–463.
Cohen, F. (1991). Trends In Computer Virus Research. Dostupné na http://vxheavens.com/lib/afc06.html.
Curtis, H. – Barnes, N. S. (1989). Biology. Worth Publishers.
Dawkins, R. ([1986] 2002). Slepý hodinář. Přel. T. Grim, Praha, Paseka.
Dawkins, R. ([1989] 2003). Sobecký gen. Podle 2. vyd. přel. V. Kopský, Praha, Mladá fronta.
Dawkins, R. (1982). The Extended Phenotype. Oxford, Oxford University Press.
Dawkins, R. (1982a). Replicators and Vehicles. R. Brandon and R. Burian (eds.). Genes, Organisms, Populations. Cambridge, Mass., The MIT Press, s. 161–180. Dostupné na: http://www.stephenjaygould.org/library/dawkins_replicators.html.
De Duve, Ch. (1995). Vital Dust. Life as a cosmic imperative. New York, BasicBooks.
Dennett, D. ([1997] 2004). Druhy myslí. K pochopení vědomí. Přeložil J. Peregrin, Praha, Acdemia.
Dennett, D. (1995). Darwin’s Dangerous Idea. New York: Simon & Schuster.
Durham, W. H. (1992). Coevolution: Genes, Culture, and Human Diversity. Stanford, Cal., Stanford University Press.
Futuyma, D. J. (1986). Evolutionary Biology. Sunderland, Mass., Sinauer Associates.
Futuyma, D. J. (2005). Evolution. Sunderland, Mass., Sinauer Associates.
Ghiselin, M. (1984). The Triumph of the Darwinian Method. The Uni. of Chicago Press, Chicago.
Gould, S. J. – Lewontin, R. (1979). The spandrels of San Marco and the Panglossian paradigm: a critique of the adaptationist programme. Proc. R. Soc. Lond., B, Biol. Sci. 205 (1161), 581–598.
Havlík, V. (2008). Ontologie evoluční perspektivy. In: J. Nosek, V. Havlík (eds.), Evoluce a věda. s. 121–141.
Havlík, V. (2011). The Scientific Status of Darwinism. In: T. Hříbek, J. Hvorecký (eds.), Knowledge, Value, Evolution. London: College Publications, King’s College, s. 85-98.
Havlík, V. (2012). Vstříc univerzálnímu evolučnímu principu. In: V. Havlík, T. Hříbek at al. Z evolučního hlediska. Pojem evoluce v současné filosofii. Filosofia, Praha.
Hendrix, R. W. et al. (1999). Evolutionary Relationships among Diverse Bacteriophages and Prophages: All the World’s a Phage. Proceedings of the National Academy of Sciences 96, s. 2192–2197.
Hodge, M. J. S. (1977) The Structure and Strategy of Darwin’s ‚Long Argument‘. British Journal for the History of Science, vol. 10, s. 237-246.
Hodge, M. J. S. (1987) Natural Selection as a Causal, Empirical and Probabilistic Theory. In: L. Krüger (ed.), The Probabilistic Revolution, vol. 2., MIT Press, Cambridge, MA 1987, s. 233-270.
Holland, J. H. (1975). Adaption in Natural and Artificial Systems: An Introductory Analysis with Applications to Biology, Control, and Artificial Intelligence. Cambridge, Mass., The MIT Press.
Hull, D. L. – Langman, R. E. – Glenn, S. S. (2001). A General Account of Selection: Biology, Immunology, and Behavior. Behavioral and Brain Sciences 24, s. 511–528.
Hull, D. L. (1988). Science as a Process: An Evolutionary Account of the Social and Conceptual Development of Science. Chicago, The University of Chicago Press.
Hull, D. L. (2001). Science and Selection. Cambridge, Cambridge University Press.
Kitcher, P. (1985) Darwin’s Achievement. In: N. Rescher (ed.), Reason and Rationality in Natural Science, Uni. press of Amerika, Lantham, s. 127-189.
Koza, J. R. (1992). Genetic Programming: On the Programming Computers by Means of Natural Selection. Cambridge, Mass., The MIT Press.
Lewontin, R. C. (1970). The units of selection. Annual Review of Ecology and Systematics, 1, s. 1–18.
Lloyd, E., (1983). The Nature of Darwin’s Support for the Theory of Natural Selection. Philosophy of Science, vol. 50, s. 112-129.
Mayr, E. (2009). Co je evoluce? Z angl. přel. Z. Urban. Praha, Academia.
Morgan, G. J. – Pitts, W. B. (2008). Evolution without Species: The Case of Mosaic Bacteriophages. British Journal for the Philosophy of Science 59, s. 745–765.
Obitko, M. (2010). Genetic Algorithms. Dostupné na http://www.obitko.com/tutorials/genetic-algorithms/ga-basic-description.php.
Oyama, S. – Griffiths, P. E. – Gray, R. D. (2001). Cycles of Contingency: Developmental systems and evolution. Cambridge, Mass., MIT Press.
Plotkin, H. C. (1993). Darwin Machines. New York, Penguin Books.
Popper, K. ([1974] 2002). Unended Quest, Routledge, 5. vyd., s. 195.
Rechenberg, I. (1971). Evolutionsstrategie – Optimierung technischer Systeme nach Prinzipien der biologischen Evolution, (PhD thesis).
Ridley, M. (2004). Evolution. 3. vyd. Oxford, Wiley-Blackwell.
Ruse, M. (1975). Charles Darwin’s Theory of Evolution: An Analysis. Journal of the History of Biology, vol. 8, no. 2, 1975, s. 219-241.
Sintonen, M., (1990). Darwin’s Long and Short Arguments. Philosophy of Science, vol. 57, no. 4, s. 677-689.
Smolin, L. (2004). Scientific alternatives to the anthropic principle. Dostupné na <arXiv:hep-th/0407213 v3>.
Sober, E. (1985). The Nature of Selection. Paperback ed., MIT.
Thagard, P. (1978). The Best Explanation: Criteria for Theory Choice. The Journal of Philosophy, 75, s. 76-92.
Toulmin, S. (1972). Human Understanding. Princeton, N. J., Princeton University Press.
Weise, T. (2009). Global Optimization Algorithms. Theory and Application. 2. vyd. Dostupné na http://www.it-weise.de/projects/book.pdf.
Williams, M. B. (1970). Deducing the consequences of evolution: A mathematical model. Journal of Theoretical Biology, 29, s. 343–385.
Wilson, F. (1992). Empiricism and Darwin’s Science. Kluwer, Dordrecht.
Zurek, W. (2003). Quantum Darwinism: Entanglement, branches, and the emergent classicality of redundantly stored quantum information. Dostupné na: <arXiv:quant-ph/0505031>.
Zpracoval: PhDr. Vladimír Havlík, CSc., Filozofický ústav AV ČR v Praze.