Paměť
Posted By 0003 On 13.7.2013 @ 7:16 In Učení a paměť | Comments Disabled
Úvod
Paměť (nervová paměť) je neurofyziologický a psychický proces, prostřednictvím kterého si uchováváme vědomosti a osvojujeme zkušenosti, které jsou nezbytné pro život. Odráží minulé prožívání a chování ve vědomí člověka. Opakem je zapomínání.
Paměť nám umožňuje vzdělání, výchovu, uchování informací. Utváří individuální zkušenost a psychický vývoj. Je součástí učení, což je vytváření dlouhodobé pamětní stopy obvykle opakovanou prezentací podnětů. Do procesu paměti patří i zapomínání.
- Paměť dodnes není vysvětlena komplexně.
- Každá teorie vysvětluje pouze určité stránky paměti.
Fyziologie paměti
Paměť je zajištěna plastičností nervové soustavy
- Nervové buňky mohou vytvářet dočasné spoje;
- Dočasné spoje jsou umožněny stopami po vzruchu v mozkových buňkách a synapsích – především v mozkové kůře a hippokampu;
- Jedná se o fyziologické změny v nervových buňkách - pro svůj vznik potřebují určitý čas na dozrání a vývoj.
Fáze paměti
Rozlišujeme následující fáze paměti:
- Vštípení (encoding, impregnace)
- Vštípení znamená uložení informací do paměti. Po fyziologieké stránce jde o vytvoření dočasných spojů v mozkové kůře působením vnitřních a zevních podnětů.
- Toto uložení může být buď úmyslné nebo neúmyslné:
- Úmyslné (záměrné) ukládání do paměti se děje především při cílevědomém učení;
- Neúmyslné (bezděčné) uložení do paměťových stop vzniká především za okolností citově podbarvených anebo jinak nápadných.
- Uchování (storage, retence)
- Doba zapamatování – čas, který uplyne od zapamatování do možného vybavení. Minulé zážitky setrvávají v naší psychice, aniž by existovaly v našem vědomí.
- K upevňování vložené informace slouží opakování (viz níže).
- Vybavování (retrieval, reprodukce) – je oživováním vytvořených mozkových stop a dočasných spojů, kdy ve vědomí se znovu objevují minulé zážitky.
- Zapomínání – je postupným (fyziologickým) vyhasínáním vytvořených mozkových stop (dočasných nervových spojení), nebo také vzniká v důsledku atrofie, postupného rozpadu nervové tkáně.
- Zapomínání znamená ztrátu přesnosti vložené informace.
- Mění se kvalita i kvantita (množství vybaveného je menší než zapamatovaného);
- Přeskupují se jednotlivé složky (mění se struktura);
- Provází je změna jistoty v přesnosti vybaveného;
- Zzmenšuje se pohotovost k vybavení zapamatovaného.
Zapomínání má pro fyziologii paměti u člověka kladný i záporný význam. Kladný v tom, že se zapomínají se nepříjemné, tragické a otřesné zážitky. Pokud by přetrvávaly, narušovaly by harmonii duševního života a zdravý vývoj osobnosti. Záporný v tom, že se zapomínají vědomosti, dovednosti, návyky, způsoby chování atd.
Proti zapomínání působí opakování, uplatňování nově nabytých poznatků v praxi, a to především těmito mechanismy:
- Reprodukcí –vybavením si totožné informace (memorování, učební dril, opakování);
- Vzpomínáním – informaci si vybavujeme na základě logických souvislostí a různých asociací, informace není totožná, ale přece přesná;
- Znovupoznáním – jestliže se s nějakým podnětem setkáme znovu, uvědomíme si, že nám už je známý.
Klasifikace paměti
V dělení paměti a výčtu jednotlivých druhů a typů paměti existuje obrovská rozmanitost. Paměť je možno dělit podle několika hledisek. Je prakticky nemožné podat jednotné klasifikační schéma paměti. Níže uvedené schéma na obrázku 1 proto v nejlepším případě je pouze jedním z příkladů.
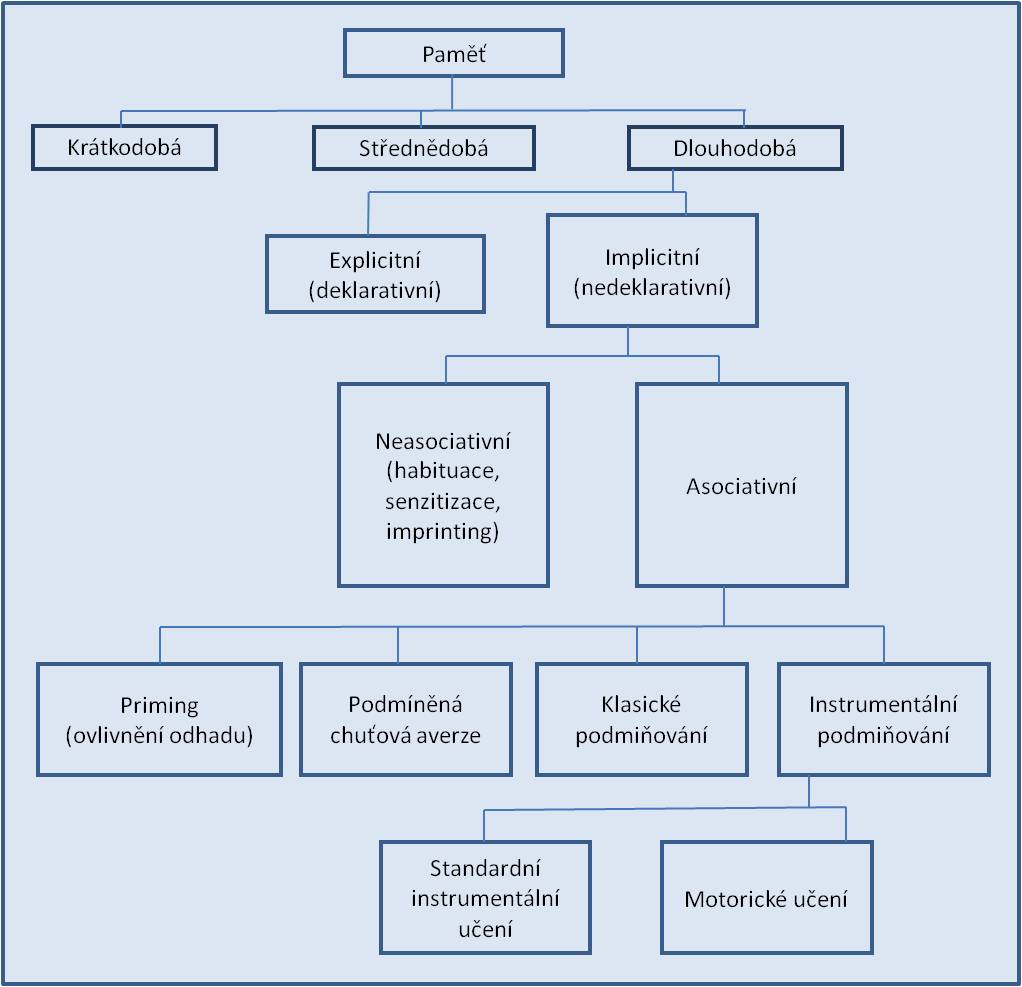 [1]
[1]
Obr. 1. Klasifikace paměti.
Druhy paměti podle délky trvání
Nejběžnější dělení paměti je na okamžitou, krátkodobou a trvalou. Je důležité rozlišovat mezi pamětí umožňující explicitní vybavení informací během krátkého intervalu po jejich uložení a pamětí, která zahrnuje delší časové rozmezí. Podrobnější dělení paměti podle délky jejího trvání je následující:
- Paměť okamžitá (ikonická anebo také echoická);
- Paměť krátkodobá („recentní“);
- Paměť střednědobá;
- Paměť dlouhodobá (trvalá, referenční, permanentní).
Okamžitá paměť
Trvá jen několik vteřin, informace se v ní udrží ve stavu, v jakém přišla, další zpracování informace – nebo její vyřazení – se teprve připravuje.
Krátkodobá paměť
Krátkodobá paměť (obr. 1, 2) trvá několik vteřin až minut (většinou do 30 – 40 sekund až minuty), můžeme ji prodloužit opakováním informací v duchu. Je omezená a uchovává se v ní jen několik informací (8 dekadických číslic, 5 jednoslabičných slov). Přechovává informace, které jsou po krátkou dobu potřebné k vykonání určité činnosti. Tyto informace lze dále zpracovávat i s nimi manipulovat. Proto zřejmě je do jisté míry také totožná s tzv. pracovní pamětí (viz níže). Zatím není jasné, jakou funkci krátkodobá paměť vlastně má. Existuje více teorií, zda je vstupní branou k paměti dlouhodobé, nebo jen jakousi její formou. Další možností je, že se jedná jen o velké množství různých momentuálně probíhajících procesů.
Krátkodobá paměť se označuje i jako primární paměť. Pokládá se za vstupný paměťový proces. Typickým příkladem je telefonní číslo, které vyčteme ze seznamu a jdeme na ně telefonovat - v paměti ho podržíme několik sekund, event . dokud se nedovoláme). Z krátkodobé paměti se přesouvají do dlouhodobé paměti jen ty informace, které si chceme nebo potřebujeme uchovat.
Můžeme tedy krátkodobou paměť považovat za jakýsi filtr, přes který přechází jen významnější podněty (např. do dlouhodobé paměti se dostanou jen čísla, na která často voláme a která nechceme zapomenout). Přesun z krátkodobé paměti do dlouhodobé paměti se nazývá konsolidace, upevnění paměti, která trvá několik minut až hodin (většina informací je konsolidována do 30 minut). Během této doby je informace labilní a dojde-li k přerušení dějů, informace se ztrácí.
Mechanismy krátkodobé paměti
Krátkodobá paměť je založena na vzájemném propojení tří funkčních okruhů - centrálního exekutivního (asociační kůra dorzolaterální frontální oblasti) a dvou daslších, zahrnujících unimodální asociační zrakové a sluchové oblasti. Krátkodobá paměť může být zachována i při rozsáhlejším postižení mediotemporální oblasti.
Pravděpodobným, i když zatím neprokázaným nervovým podkladem krátkodobé paměti jsou reverberační nervové obvody anebo pozitivní zpětnovazební okruhy. Jejich podstatou jsou zpětná synaptická spojení postsynaptických neuronů s presynaptickými neurony zapojenými v sérii, případně i zpětná synaptická spojení z kolaterál axonu neuronu na tělo anebo dendrity téhož neuronu. Nervové vzruchy potom cirkulují v uzavřeném kruhu, takže dochází k opakované re-excitaci neuronů generujících akční potenciály. Účinnost tohoto mechanismu krátkodobé paměti dokazuje i dráždění korových neuronů elektrickými podněty opakovanými s určitou frekvencí – excitovaný neuron generuje nervové vzruchy ještě určitý čas po skončení dráždění.
Dlouhodobá paměť
V dlouhodobé paměti (obr. 1,2) se uchovávají informace různě dlouho - několik dní, roků, desetiletí, i celý život, jestliže se příjem informace spojoval se silným emotivním zážitkem. Informace by z této paměti neměly nikdy zmizet, časem se ale mohou stát hůře dostupnými, nebo se přepíšou informacemi novými.
Paměť explicitní a implicitní
Dlouhodobou paměť lze dále rozdělit na explicitní a implicitní (schéma 1):
- Explicitní (deklarativní) paměť se vztahuje k vědomému vybavování informací – událostí anebo faktických znalostí. Dělí se na paměť epizodickou a sémantickou:
- Epizodická paměť je zaplněna událostmi a ději, které jsme prožili dříve a nejen v krátké nejbližší době – např. co jsme dělali včera, ale také ději, ke kterým často máme osobní vztah (školní a mladá léta, citové vztahy).
- Sémantická paměť uchovává slovní a pojmové vědomosti o okolním světě – např. všeobecné deklarativní informace, pojmy, zeměpisné znalosti, matematické vzorce, rovnice a také další řadu fakt, které si osvojujeme během života.
- Implicitní (nedeklarativní) paměť ovlivňuje náš život, chování a motorické dovednosti tím, že do nich promítá naše nabyté zkušenosti bez podmínky vědomého vybavování informací („nevědomá“ paměť). Má co dělat s motorickými návyky, zručností, ale i s tvorbou percepčních schémat. Dělí se na paměť procedurální a na podněcování paměti (priming):
- Procedurální paměť nám umožní to, že opakování určité činnosti usnadní a hlavně zautomatizuje nové dovednosti v budoucím životě (chůzi, plavání, jízdu na kole, hru na hudební nástroj, ale také mluvení, psaní atd.). Pro procedurální paměť ( srovnáme-li ji s deklarativní) je typické:
- Pomalé učení;
- Postupné zdokonalování výkonu;
- Výrazná odolnost proti zapomínání.
- Podněcování (priming) je zvýšení paměťové výkonnosti záhy poté, kdy jsme byli vystaveni podnětům, které měly situační vztah k danému úkolu (verbální nebo obrazové podněty).
- Procedurální paměť nám umožní to, že opakování určité činnosti usnadní a hlavně zautomatizuje nové dovednosti v budoucím životě (chůzi, plavání, jízdu na kole, hru na hudební nástroj, ale také mluvení, psaní atd.). Pro procedurální paměť ( srovnáme-li ji s deklarativní) je typické:
Implicitní (nedeklarativní) paměť se upevňuje dvěma typy učení (ve schématu neukázáno). Prvním je motorické učení, kdy je cílem vypracování nové koordinace činnosti různých svalových skupin (např. učení se chůzi, jízdě na kole anebo také mluvení a psaní). Druhým je senzorimotorická koordinace, kdy se daný člověk učí pohybovat rukou opačně než ukazuje zraková informace (např. obtáhnout obrazec, který vidí pouze v zrcadle). Deklarativní pamětní stopa může být stálým opakováním přeměněna na nedeklarativní (zaautomatizování projevu, např. automatická slovní anebo větná spojení apod.).
Asociativní a neasociativní paměť
Spojování určitého podnětu pro učení s dalšími vlivy vedlo k rozdělení paměti na asociativní a neasociativní (schéma 1).
- Při asociativním učení si organismus zapisuje vztah jednoho podnětu ke druhému. Typickým příkladem je podmíněný reflex.
- Pod neasociativní paměť spadá habituace a senzitizace.
- Habituace je vypracování návyku, útlumu, ignorace neutrálního podnětu, který se často opakuje a nemá signální význam. Částečně byl poznán i molekulární mechanismus habituace – neustálým opakováním impulsů, přicházejících na neuron, dochází ke snížení koncentrace vápenatých iontů na presynaptickém zakončení, a tím k omezení množství mediátoru uvolňovaného do synaptické štěrbiny.
- Senzitizace je naopak jev, kdy opakovaný podnět vyvolá silnější odpověď, jestliže je několikrát spojen s příjemným anebo nepříjemným pocitem. Podkladem senzitizace je prodloužení akčního potenciálu. Toho se dosáhne zvýšením obsahu Ca2+ v presynaptických zakončeních přicházejícími impulsy. Mění se konformace adenylátcyklázy, zvýšeně se tvoří cAMP, a to vede ke zvýšenému uvolňování mediátorů exocytózou do synaptické štěrbiny. Celý tento jev nazýváme presynaptická potenciace.
Mechanismy dlouhodobé paměti
Podstatou uložení paměťové informace je neuronální změna. Nazýváme ji paměťová stopa anebo paměťový engram. Dlouhodobá paměť se uchovává i po závažných poraněních mozku anebo po elktrošocích - tedy pokud nejsou zničeny mozkové struktury, které jsou za uchovávání dlouhodobých paměťových stop odpovědné. Proto se odůvodněně předpokládá, že dlouhodobé uchování paměťových stop může mít biochemický podklad. Existuje několik výkladů vytváření dlouhodobých paměťových stop.
- Jednou z možností je vytváření nových synaptických spojení.
- Jinou možností jsou funkční změny v existujících neuronálních drahách. V této souvislosti se pozornost soustřeďuje hlavně na dva mechanismy. Prvním je posttetanická potenciace, druhým je dlouhodobá (long-term) potenciace (LTP).
- Posttetanická potenciace (facilitace synaptického přenosu) vzniká jako následek hromadění Ca2+ v presynaptických zakončeních po krátké salvě impulsů presynaptického neuronu, až jsou zaplněna vazebná místa receptorů, která udržující klidovou nízkou hladinu volných iontů intracelulárního kalcia. Trvá minuty až hodiny.
- Dlouhodobá potenciace (LTP) byla pozorována především na glutámátergních zakončeních v hippokampu. Může trvat až několik dnů. Podkladem opět je vzestup intracelulárního Ca2+ v postsynaptickém neuronu.
Glutamátergní synapse
Rozlišujeme tři druhy glutamátergních synapsí. Jsou pojmenovány podle farmakologicky účinných sloučenin, které vyvolávají jejich excitaci – 1. kainátové, 2. quisqualátové (souhrnně non-NMDA-receptory) a 3. N-metyl-D-aspartátové (NMDA-) receptory. Non-NMDA-receptory způsobují krátkodobou depolarizaci přechodným zvýšením propustnosti membrán pro sodné a draselné ionty. NMDA-receptory naproti tomu jsou za klidového membránového potenciálu zablokovány hořečnatými ionty, k jejichž uvolnění dochází právě při depolarizaci.
Přicházejí-li vzruchy s frekvencí vyšší než 10 Hz, aktivace NMDA-receptorů se po příchodu následujícího impulzu časově překryje s depolarizací způsobenou otevřením non-NMDA-kanálů, a NMDA kanály se otevřou. Při frekvenci podnětů menší než 10Hz se tyto dva děje vůbec nepřekrývají. Otevření NMDA-kanálů má za následek vstup iontů Ca2+ do buňky. Ca2+-kalmodulin-dependentní kináza aktivuje proteinkinázy, které fosforylují různé bílkoviny. Dochází ke změně jejich vlastností, což má za následek zvýšení účinnosti příslušných synapsí, a to buď přetrvávající biochemickou změnou (ovlivněním propustnosti některých kanálů, vyšším množstvím uvolněného mediátoru, větším množstvím receptorů, atd.), nebo morfologickou změnou (přestavbou synapsí anebo i celých dendritů s větším množstvím bílkovin). Fosforylace některých enzymů se uplatňuje i v metabolismu kyseliny arachidonové. Její metabolity, doplňované dalším mediátorem, NO, působí zpětnovazebně – po aferentní stimulaci jsou schopné vyvolat vylití většího množství glutamátu do synaptické štěrbiny, a tím další zvýšení dráždění příslušného neuronu.
Zatím není známo, jakým způsobem se inforace zapisuje v oblastech, kde jsou NMDA-receptory zastoupeny jen v malé koncentraci. Stále se zkoumají další souvislosti – zda jsou paměťové stopy difúzní, nebo lokalizované, které části mozku se na jejich zápisu podílejí apod.
Paměť pracovní a referenční
Podle etologického hlediska můžeme paměť rozdělit na pracovní a referenční (obr. 2).
- Pracovní paměť je přechodná, střednědobá, a nepoužije-li se informace během určité doby, ztrácí význam a mizí. Přinejmenším zčásti se překrývá s krátkodobou pamětí.
- Referenční paměť je paměť trvalá, analogická dlouhodobé paměti.
 [2]
[2]
Obr. 2. Klasifikace paměti s důrazem na délku trvání. (Zdroj: Miloslav Kopeček, Kognitivní funkce – teorie a praxe; Paměť [3]).
Senzorická paměť
Některá dělení ještě odlišují tzv. senzorickou paměť (ve schématu 1 neukázáno). Senzorická paměť zahrnuje paměť vizuální, sluchovou, hmatovou atd. Informace se v každé z nich uchovávají jen určitou krátkou dobu.
- Světelná senzorická paměť
- Délka světelné senzorické paměti se liší od sluchové. Světelný vjem trvá pouze jednu desetinu sekundy (tzv. setrvačnost zrakového vjemu). Můžeme se o tom přesvědčit pokusem, kdy budeme v zatemněné místnosti sledovat pohyb libovolného bodového zdroje světla – budeme vnímat nejenom momentální polohu zdroje světla, ale i dráhu, kterou zdroj urazil během poslední desetiny sekundy.
- Sluchová senzorická paměť
- Na rozdíl od zrakového vjemu je sluchová senzorická paměť trvalejší, záznam je trvalejší (v průměru jsou to 3 sekundy), ale trvání vjemu individuálně dosti kolísá.
Typy paměti podle převládajících charakteristik paměťových stop
- Názorný typ paměti.
- Při zapamatování a vybavování se výrazněji uplatňuje první signální soustava (představy). Jedinec s tímto typem paměti si rychleji a lépe zapamatuje (vybavuje) názorné podněty.
- Slovně logický typ paměti.
- Při zapamatování a vybavování se výrazněji uplatňuje druhá signální soustava ( slova). Člověk s tímto typem paměti si rychleji a lépe zapamatuje pojmy, soudy, úsudky a myšlenky.
- Emocionální (citový) typ paměti.
- Projevuje se lehčím a rychlejším zapamatováním citů a citově zabarvených zážitků (pozitivních i negativních). Převážně to platí u malých dětí.
Anatomické struktury odpovědné za paměť
Paměť vázaná na hippokampální okruhy
Klíčovou strukturou pro správné fungování explicitní (deklarativní) epizodické paměti je orbitální kůra a mediální temporální oblast mozkové kůry. Intaktní dlouhodobá paměť je proto podmíněna neporušeným parenchymem a vzájemnými propojeními hippokampu, entorinální a peririnální kůry a kůry parahippokampálního závitu. Při izolovaném postižení hippokampů se rozvine anterográdní amnézie, zatímco pro vznik antero- i retrográdní amnézie ještě musí dojít k postižení přilehlých mediálních temporálních oblastí. I v takových případech však bývá zachována paměť na vzdálenější minulost. Ukazuje se, že vzpomínky na velmi dávné události jsou uchovávány jinde, nejspíše v zadních temporo-parietálních oblastech.
Branou deklarativní paměti je hippokampus. Všechna data, která jsou zachycena v různých korových centrech (např. zrakové či sluchové vjemy, řeč apod.) a následně uvědomována, se dostávají do hippokampální krajiny, kde jsou zpracována a dále převedena k uložení do příslušných asociačních oblastí mozkové kůry. Uchovávání informací tedy víceméně probíhá ve stejných oblastech mozku jako vnímání. Důležité také je vzájemné propojení těchto oblastí s dalšími oblastmi (reciproční kortiko-kortikální spoje) , intaktní Papezův okruh (hippocampus, corpus mamillare, mediální dorzální talamická jádra) a zachovaná cholinergní transmise (nucleus basalis Meynerti). Emočních paměťových reakcí se účastní další struktury limbického systému (amygdala, inzula atd.).
Poškození explicitní sémantické paměti není omezeno jen na mediální temporální oblast. K jejímu poškození také dochází při rozsáhlých lézích kůry temporální, parietální a částečně i okcipitální oblasti. Typickým příkladem např. může být herpetická encefalitida nebo degenerativní demence (Alzheimerova nemoc nebo frontotemporální demence).
Paměť nevázaná na hippokampální okruhy
Implicitní („nevědomá“) paměť není vázána na hippokampální okruhy. Při řízení auta provádíme jednotlivé cílené pohyby automaticky, aniž bychom museli pokaždé cíleně přemýšlet, jak sešlápnout spojku a správně zařadit rychlost. Implicitní procedurální paměť se realizuje v úzké vazbě na subkortikální motorické okruhy, zejména mozečku a bazálních ganglií. Okruhy začínají v senzorické kůře, postupují do bazálních ganglií a do mozečku, odtud zpět do ventrálního talamu a do premotorické kůry. Implicitní paměť proto není porušena při mediotemporálních afekcích, ale může být ovlivněna dopaminergními látkami.
Neasociační učení využívá reflexní dráhy.
Kazuistika
- Tyto znalosti byly doplněny velmi názornými kazuistikami pacientů s poškozením určité části mozku. Často se uvádí ilustrativní případ pcienta H.M., který trpěl selektivním postižením deklarativní paměti. Ve věku devíti let utrpěl pádem z kola kraniocerebrální trauma a postupně se u něj rozvinula sekundární epilepsie rezistentní na léčbu. Proto bylo přikročeno k operativnímu odstranění hippokampů a přilehlých spánkových oblastí. Od té doby přestaly záchvaty, ale vzniklo trvalé poškození paměti.
- Pacient si dobře pamatoval na události, které předcházely jeho operaci. Nové informace ale dokáže podržet jen po velmi krátkou dobu, není schopen je uložit do dlouhodobé paměti. Například za určitou dobu po jídle zapomene, že už jedl, a přeje si pokračovat v jídle; nebo poznává sám sebe na fotografiích z dětství, ale už ne při pohledu do zrcadla. Implicitní (dlouhodobá) paměť (podněcování, priming) je poměrně zachována, takže když si například přečetl určité slovo a po několika hodinách mu bylo předloženo totéž slovo v neúplné podobě s chyběním některých písmen, byl schopen ho „rozluštit“ podstatně rychleji než jiná slova, se kterými se před tím nesetkal; při tom si ale vůbec neuvědomoval, že dané slovo už před chvílí spatřil.
Vlastnosti a individuální zvláštnosti paměti
Vlastnostmi paměti nazýváme individuální rozdíly v kvalitě paměti. Patří k nim:
- Rychlost zapamatování;
- Pohotovost vybavování;
- Šíře paměti;
- Přesnost zapamatování a vybavování
- Trvání pamatování (délka udržení v paměti).
Cvičení (trénování) paměti
- Opakováním naučeného;
- Prohlubováním paměťových stop;
- Zvyšováním pozornosti, soustředěním;
- Logickým vysvětlením naučeného, odvozováním;
- Vysvětlováním druhému;
- Zapisováním;
- Používáním mnemotechnických pomůcek;
- Motivací (odměna, trest).
Ontogenetický vývoj paměti
- Prenatální období:
- Plod reaguje na podněty, zvuky (akustické – zvuk se dobře šíří tělními tekutinami a plodovou vodou, proto reakce na hudbu, která se mu líbí, na matčin zpěv, má schopnost sociální interakce s matkou).
- Novorozenecké období:
- Novorozenec si pamatuje hlas matky z prenatálního období, dominují nepodmíněné reflexy (sací, polykací, vylučovací atd.), reaguje na podněty , má pouze příjemné anebo nepříjemné pocity, vnímá je nediferencovaně celým tělem.
- Kojenecké období:
- Okolo 6. měsíce se objevuje možnost uchování stop v paměti po dobu 2 – 3 dnů;
- Okolo 1. roku možnost uchování stop v paměti v rozsahu 2 – 3 týdnů;
- Strach z neznámého (i osob), objevuje se schopnost předvídat následky činnosti (naučí se vynucovat si pozornost okolí).
- Batolecí období:
- Citovost, bezděčnost, obraznost (konkrétní jevy), mechanické zapamatování, vybavuje si pomocí znovupoznání.
- Předškolní věk:
- Citovost, mechanická paměť, ke konci období i paměť slovně logická.
- Mladší školní věk:
- Záměrná paměť, začíná převažovat logická paměť.
- Puberta:
- Logická paměť, v učení jsou velké výkyvy.
- Adolescence:
- Záměrná paměť, převažuje logická paměť.
- Dospělost:
- Mladší dospělost – paměť velmi výkonná;
- Střední dospělost – paměť mechanická, zlepšuje se slovně-logická;
- Starší dospělost – paměť slábne, ztrácí rychlost, pružnost, vynalézavost, proces učení se zpomaluje.
- Stáří
- Paměť se oslabuje, převládá hlavně mechanická paměť, snadné zapomínání běžných věcí a událostí, zejména čerstvých.
Použitá literatura a literatura k dalšímu studiu
Ganong, W. F.: Přehled lékařské fyziologie, H+H, Jinočany 1999.
Gembalová I.: Mechanismy paměti, 2003 [4].
Jirák R.: Poruchy paměti – základní přehled.: Medicína po promoci 6/2011.
Rusina R.: Paměť a její poruchy, Neurologie pro praxi 4/2004.
Zpracovala: doc. MUDr. Květoslava Dostálová, CSc., Ústav patologické fyziologie LF UP v Olomouci.
Article printed from E-learningová podpora mezioborové integrace výuky tématu vědomí na UP Olomouc: http://pfyziolmysl.upol.cz
URL to article: http://pfyziolmysl.upol.cz/?p=4495
URLs in this post:
[1] Image: http://pfyziolmysl.upol.cz/wp-content/uploads/2013/02/SchePamet.jpg
[2] Image: http://pfyziolmysl.upol.cz/wp-content/uploads/2013/07/ObrPamet1.jpg
[3] Miloslav Kopeček, Kognitivní funkce – teorie a praxe; Paměť: http://www.miloslavkopecek.cz/kognice.htm
[4] Mechanismy paměti, 2003: http://fyziologie.lf2.cuni.cz/uceni/lecture_notes/mechanismy_pameti/index.htm
Click here to print.